Репликация днк в цикле

Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 29 апреля 2021; проверки требуют 2 правки.
У этого термина существуют и другие значения, см. Репликация.
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК-полимераза (Polα), (4) ДНК-лигаза, (5) РНК-праймер, (6) праймаза, (7) фрагмент Оказаки, (8) ДНК-полимераза (Polδ), (9) хеликаза, (10) белки, связывающие одноцепочечную ДНК, (11) топоизомераза
Процесс репликации: раскручивание двойной спирали ДНК – синтез комплементарных цепей ДНК-полимеразой – образование двух молекул ДНК из одной
Реплика́ция (от лат. replicatio – возобновление) – процесс создания двух дочерних молекул ДНК на основе родительской молекулы ДНК. Репликацию ДНК осуществляет сложный комплекс, состоящий из 15-20 различных белков-ферментов, называемый реплисомой (англ.)[1]. С помощью специальных ферментов двойная спираль материнской ДНК расплетается на две нити, на каждой образовавшейся нити достраивается вторая нить, образуя две идентичных дочерних молекулы ДНК, которые затем скручиваются в отдельные спирали. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение.
История изучения[править | править код]
Каждая молекула ДНК состоит из одной цепи исходной родительской молекулы и одной вновь синтезированной цепи. Такой механизм репликации называется полуконсервативным. В настоящее время этот механизм считается доказанным благодаря опытам Мэтью Мезельсона и Франклина Сталя (1958 г.)[2]. Ранее существовали и две другие модели: «консервативная» – в результате репликации образуется одна молекула ДНК, состоящая только из родительских цепей, и одна, состоящая только из дочерних цепей; «дисперсионная» – все получившиеся в результате репликации молекулы ДНК состоят из цепей, одни участки которых вновь синтезированы, а другие взяты из родительской молекулы ДНК. Молекула ДНК разрезается пополам и образуются два шаблона. Два шаблона выходят из репликационной вилки. Если представить их в выпрямленном виде, то можно видеть линейку из гребёнок, которые соединены концами, но имеют промежуток. Представим, что одна гребёнка синяя, а другая – красная. Теперь подставим нижнюю красную (она из пяти гребней, как и верхняя) пятым концом к третьему верхнему (третьей верхней иголке). Удлиним цепь и сверху, и снизу. Как бы получится: пять, три, пять и т. д.- наверху и снизу тоже. Потом к этим гребёнкам добавляются после выхода шаблонов (гребёнок) из репликационной вилки ещё два шаблона. Из одной молекулы ДНК получается две идентичные материнской (если нет мутаций) молекулы, это называется полуконсервативностью.
Общие представления[править | править код]
Репликация ДНК – ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
- инициация репликации
- элонгация
- терминация репликации.
Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятие репликон. Репликон – это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий[1].
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка – место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации в электронный микроскоп можно наблюдать репликационный глазок – участок хромосомы, где ДНК уже реплицирована, окружённый более протяжёнными участками нереплицированной ДНК[1].
В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500-5000 – у эукариот[3].
Ферменты и их функции
| Фермент | Функция |
|---|---|
| ДНК-гираза | Относится к группе топоизомераз. Вносит временные двуцепочечные разрывы в ДНК, облегчая её разматывание. |
| Хеликаза | Разделяет цепи двухцепочечной молекулы ДНК на одинарные цепи. |
| SSB-белки | Связывают одноцепочечные фрагменты ДНК и предотвращают комплементарное спаривание. |
| Праймаза | Синтезирует РНК-затравку (праймер) – короткий фрагмент РНК, которая является инициатором в работе ДНК-полимеразы (полимераза не способна синтезировать ДНК с нуля, но может добавлять нуклеотиды к уже имеющимся). |
| ДНК-полимераза | Синтезирует ДНК, связываясь с праймером. Следует отметить, что один конец материнской ДНК полимераза синтезировала непрерывно и в одном направлении, а второй – в противоположном – фрагментами. |
| Белки скользящего зажима (застежки) | Окружают кольцом ДНК и «скользят» по ней вместе с продвигающейся вперед фермента ДНК-полимеразы. Они предотвращают диссоциацию фермента от матрицы ДНК и повышают эффективность его работы. |
| РНКаза H | Удаляет уже ненужные фрагменты РНК-затравки. |
| ДНК-лигаза | Сшивает фрагменты ДНК (фрагменты Оказаки). |
| Теломераза | Добавляет особые повторяющиеся последовательности нуклеотидов к одному концу цепи ДНК на участках теломер, тем самым компенсируя их укорачивание во время деления. |
| Реплисома (комплекс всех ферментов репликации) | Движется вдоль молекулы ДНК-матрицы, расплетая её и наращивая комплементарные цепи ДНК. |
Молекулярный механизм репликации[править | править код]
Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у прокариот[уточнить] осуществляется несколькими разными ДНК-полимеразами. ДНК-полимераза I действует на запаздывающей цепи для удаления РНК-праймеров и дорепликации очищенных мест ДНК. ДНК полимераза III – основной фермент репликации ДНК, осуществляющий синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе запаздывающей цепи. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Характеристики процесса репликации
- матричный – последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности;
- полуконсервативный – одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая – материнской;
- идёт в направлении от 5′-конца новой молекулы к 3′-концу;
- полунепрерывный – одна из цепей ДНК синтезируется непрерывно, а вторая – в виде набора отдельных коротких фрагментов (фрагментов Оказаки);
- начинается с определённых участков ДНК, которые называются сайтами инициации репликации (англ. origin)[4].
Примечания[править | править код]
Литература[править | править код]
- Сохранение ДНК в ряду поколений: Репликация ДНК (Фаворова О.О., СОЖ, 1996)PDF (151 KB)
- Репликация ДНК (анимация) (англ.)
Источник
Репликация ДНК. Вспомним, что удвоение молекул ДНК, которое осуществляется в синтетическом (S) периоде клеточного цикла, называется репликацией. *После открытия Дж. Уотсоном и Ф. Криком структуры молекулы ДНК было выдвинуто несколько гипотез о возможном механизме протекания этого процесса (рис. 16.2).
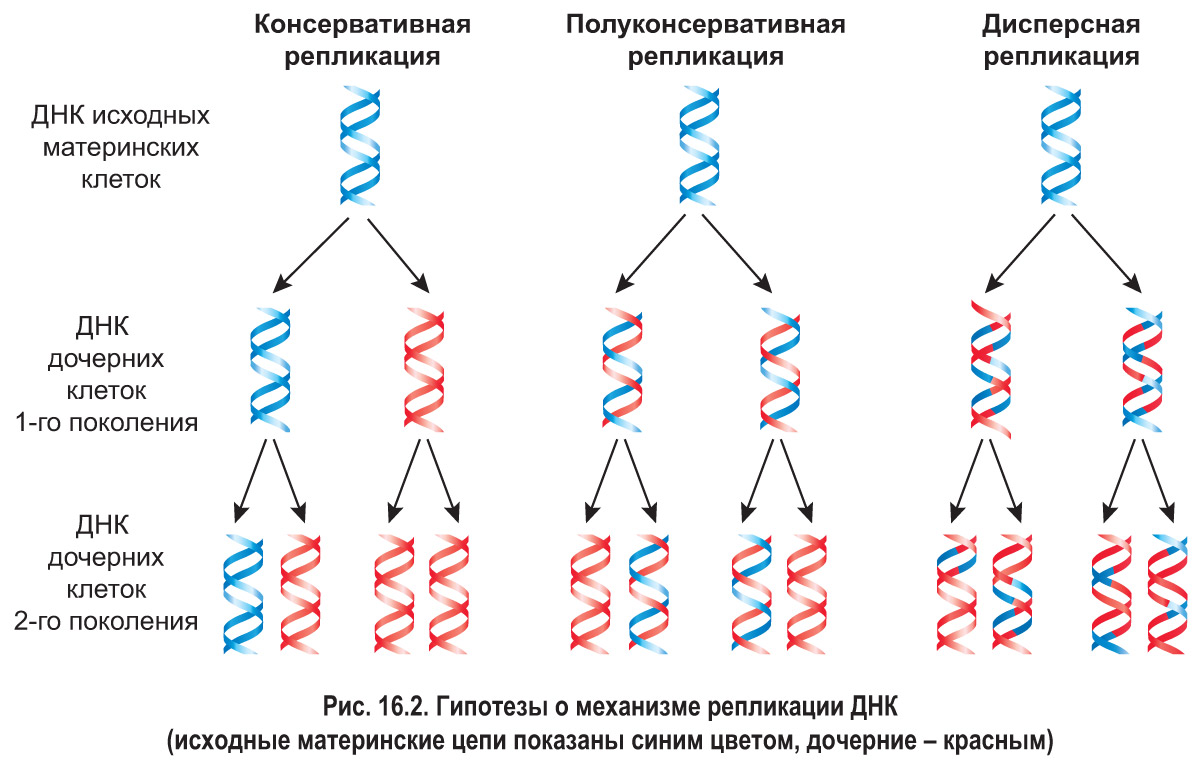
Согласно гипотезе консервативной репликации двухцепочечная материнская молекула ДНК как единое целое служит матрицей для образования дочерней молекулы, состоящей из двух совершенно новых цепей. Полуконсервативный механизм репликации предполагает разделение цепей исходной молекулы ДНК. При этом каждая материнская цепь является матрицей для синтеза дочерней цепи. Таким образом, при репликации образуются две молекулы ДНК, каждая из которых включает одну материнскую цепь и одну новую – дочернюю. Гипотеза дисперсной репликации состояла в том, что материнская ДНК распадается на фрагменты, которые выступают в роли матриц для построения отдельных участков новых молекул. Образованные таким способом молекулы ДНК должны состоять из чередующихся фрагментов исходной молекулы и вновь синтезированных.
В 1958 г. американские биологи М. Мезельсон и Ф. Сталь провели экспериментальную проверку этих гипотез. На протяжении нескольких поколений они выращивали бактерии (а именно кишечную палочку) в питательной среде, содержащей «тяжелый» азот 15N. За это время 15N вошел в состав бактериальных молекул ДНК. Далее бактерии были перенесены в среду, содержащую изотоп 14N. Следовательно, в состав вновь синтезированных цепей ДНК включался уже «легкий» азот. Из бактериальных клеток новых поколений, образовавшихся в среде с 14N, выделяли ДНК и центрифугировали в градиенте плотности хлорида цезия (рис. 16.3).

Выяснилось, что ДНК дочерних клеток первого поколения имела плотность, среднюю между плотностью «легкой» ДНК, содержащей только 14N, и «тяжелой», включающей только 15N. То есть такая «гибридная» ДНК содержала одновременно 14N и 15N. Это противоречило гипотезе о консервативном механизме репликации, согласно которой ДНК должна была разделиться на две фракции – «легкую» и «тяжелую».
ДНК, выделенная из клеток второго поколения, разделялась на «легкую» и «гибридную». Этот факт полностью соответствовал гипотезе полуконсервативной репликации и позволил исключить дисперсный механизм удвоения ДНК, согласно которому плотность ДНК второго поколения бактерий должна быть средней между «легкой» и «гибридной». Таким образом был доказан полуконсервативный механизм репликации ДНК.*
*К настоящему времени установлено, что в осуществлении репликации принимает участие целый комплекс ферментов. Также известно, что этот процесс начинается в строго определенных участках молекулы ДНК – так называемых точках начала репликации. Бактериальная хромосома, как правило, имеет одну такую точку. У ядерных организмов каждая молекула ДНК (хромосома) содержит множество точек начала репликации. Следовательно, у эукариот процесс репликации одновременно протекает на многих участках одной и той же хромосомы. Это значительно сокращает время удвоения молекул ДНК.
Процесс репликации ДНК подразделяют на три этапа: инициацию, элонгацию и терминацию.
1. Инициация (запуск). Особые ферменты начинают раскручивать молекулу ДНК от точки начала репликации. Затем фермент хеликаза разъединяет цепи исходной материнской молекулы, разрывая водородные связи между комплементарными азотистыми основаниями. При этом две цепи расходятся под определенным углом и образуют так называемую репликативную вилку, напоминающую букву Y (рис. 16.4). Хеликаза, подобно бегунку застежки «молния», движется вдоль молекулы ДНК, разделяя комплементарные цепи. Это сопровождается перемещением репликативной вилки и, образно говоря, расстегиванием «молнии». Далее с разошедшимися цепями связываются молекулы ДНК-полимеразы – главного фермента репликации.
2. Элонгация (удлинение, наращивание дочерних цепей ДНК). Молекулы ДНК-полимеразы начинают двигаться вдоль материнских цепей, используя их в качестве матриц для построения новых дочерних цепей. Это значит, что в растущие дочерние цепи ДНК включаются только те нуклеотиды, которые комплементарны соответствующим нуклеотидам материнских цепей. Поэтому репликацию относят к реакциям матричного синтеза.
Следует отметить, что в процессе репликации материалом для синтеза дочерних цепей ДНК являются не нуклеотиды как таковые, а нуклеозидтрифосфаты. От обычных нуклеотидов их отличает наличие трех остатков фосфорной кислоты (как в молекуле АТФ) вместо одного. Связи между этими остатками являются макроэргическими. Таким образом, нуклеозидтрифосфаты – соединения, богатые энергией. В процессе их присоединения к растущим дочерним цепям ДНК происходит отщепление двух «лишних» остатков фосфорной кислоты. В результате выделяется энергия, которая используется для протекания реакции (т. е. для образования фосфоэфирных связей), и нуклеозидтрифосфаты становятся стандартными нуклеотидами новых цепей ДНК.
Особенность ДНК-полимеразы состоит в том, что она может двигаться вдоль существующей цепи ДНК только в направлении 3′ → 5′. При этом наращивание дочерней цепи всегда происходит антипараллельно: от 5′-конца к 3′-концу (т. е. новые нуклеотиды добавляются к 3′-концу синтезируемой цепи). Например, если участок материнской цепи ДНК содержит последовательность нуклеотидов 3’ГТАЦАГ5′, то при наращивании соответствующего ему участка дочерней цепи нуклеотиды будут присоединяться в следующем порядке: 5’ЦАТГТЦ3′.
Способность ДНК-полимеразы к перемещению вдоль цепи ДНК только в одном направлении приводит к следующему. Вдоль одной материнской цепи (той, у которой направление 3′ → 5′ совпадает с направлением перемещения репликативной вилки) фермент движется непрерывно. Дочерняя цепь ДНК, которая при этом синтезируется, называется лидирующей или ведущей (рис. 16.5).
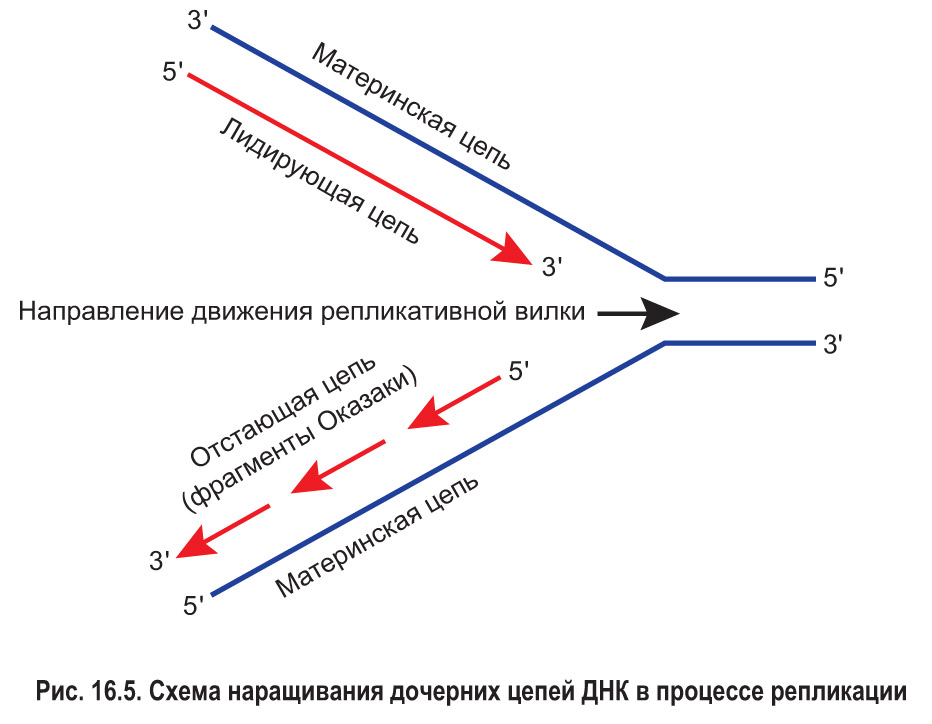
В то же время движение ДНК-полимеразы, которая работает над созданием другой дочерней цепи, не может быть непрерывным. По мере смещения репликативной вилки фермент «забегает вперед» и затем, возвращаясь назад, синтезирует новую цепь отдельными фрагментами. В честь первооткрывателей – супругов Р. и Ц. Оказáки – эти участки называют фрагментами Оказаки. Таким образом, вторая дочерняя цепь строится прерывисто с существенным отставанием от лидирующей цепи. Поэтому данная цепь получила название отстающей или запаздывающей. Впоследствии фрагменты отстающей цепи сшивает друг с другом фермент ДНК-лигаза.*
*Открытие фермента, катализирующего биосинтез ДНК, принадлежит американскому биохимику А. Корнбергу. Энзим, выделенный из клеток кишечной палочки, ученый назвал ДНК-полимеразой. В 1957 г. с помощью ДНК-полимеразы А. Корнберг впервые осуществил синтез ДНК в лабораторных условиях, а в 1959 г. за открытие механизмов биосинтеза нуклеиновых кислот он совместно с биохимиком С. Очоа был удостоен Нобелевской премии.*
*3. Терминация (остановка). Когда репликативная вилка достигает соседнего участка ДНК, на котором также осуществлялась репликация, ферменты завершают свою работу.* В результате репликации образуются две молекулы ДНК, идентичные друг другу и исходной материнской молекуле. В состав каждой из них входит одна цепь материнской молекулы ДНК и одна вновь синтезированная дочерняя цепь.
*Каждая клетка существует определенное время, по истечении которого либо делится, либо погибает. Различают два типа клеточной гибели – некроз и апоптоз.
Некроз – это гибель клеток, обусловленная действием повреждающих факторов. Причинами некроза может быть воздействие на клетки высоких или низких температур, ионизирующих излучений, токсических веществ, в том числе выделяемых болезнетворными микроорганизмами. Некротическая гибель клеток наблюдается также в результате их механического повреждения, нарушения иннервации и кровоснабжения тканей, в состав которых они входят, и т. д.
При некрозе обычно происходит гибель не отдельных клеток, а целых их групп. Например, в результате ожога массово погибают клетки кожи, подвергшиеся действию высокой температуры. При инфаркте миокарда из-за прекращения снабжения кислородом отмирает участок сердечной мышцы, состоящий из множества клеток. В поврежденных клетках нарушается проницаемость плазмалеммы и внутриклеточных мембран, прекращаются синтез белков и другие процессы метаболизма, происходит разрушение ядра и органоидов. Клетки погибают, и продукты их распада, как правило, стимулируют запуск воспалительной реакции в зоне некроза.
Апоптоз, который иногда называют клеточным самоубийством, представляет собой запрограммированную гибель клеток, которая регулируется организмом. Этот процесс обнаружен у различных групп живых организмов, в том числе и одноклеточных. У животных и растений апоптоз наблюдается на всех этапах индивидуального развития, даже в эмбриональный период. Известно, например, что в организме взрослого человека ежедневно в результате апоптоза запланированно погибает 50-70 млрд клеток.
Уничтожение клетки путем апоптоза могут запускать как определенные внеклеточные сигналы, так и внутриклеточные факторы. В результате апоптоза клетка распадается на отдельные фрагменты, окруженные плазмалеммой. Затем эти фрагменты поглощаются лейкоцитами или соседними клетками без запуска воспалительной реакции.
Благодаря апоптозу в организме поддерживается клеточный гомеостазис, т. е. определенное соотношение между различными типами клеток. Апоптоз играет важную роль в процессах клеточной дифференцировки, необходимой для правильного развития и функционирования тканей и органов. Он также обеспечивает уничтожение различных дефектных клеток, например мутантных. За открытие механизмов апоптоза и генетической регуляции развития органов британские биологи С. Бреннер и Дж. Салстон, а также американский ученый Р. Хорвиц в 2002 г. были удостоены Нобелевской премии.*
Источник