Цикл развития листостебельного мха протонема

Мхи – отдел высших растений, преимущественно многолетних. Отличается регрессивным развитием спорофита, в жизненном цикле мхов доминирует (преобладает) гаметофит. Мхи изучает бриология (от греч. bryon – мох, и logos – слово) – наука о мхах.
Эволюционно мхи представляют тупиковую (слепую) ветвь развития растений, во многом близки к водорослям. Для размножения большинству мхов нужна вода, так как мужские половые клетки могут добраться до женских только с ее помощью. Мы рассмотрим этот отдел на примере кукушкина льна, который относится к подклассу Зеленые мхи.
Интересный факт – устьица у мхов отсутствуют, вместо них имеются поры без замыкающих клеток. У моховидных отсутствуют настоящие стебли и листья, которые мы привыкли видеть у остальных высших растений, нет корней и специализированных проводящих тканей. Механические ткани развиты слабо, так что по совокупности причин рост в высоту для мхов оказывается затруднительным (ограниченным).

Строение кукушкина льна
Кукушкин лен относится к двудомным растениям. Стебель у него прямостоячий, это один из наиболее высокорослых мхов, достигающих в высоту 50 см и более. Отсутствие корней компенсируют нитевидные выросты – ризоиды, которые обеспечивают прикрепление растения к субстрату, всасывают воду и растворенные в ней минеральные вещества (в отличие от ризоидов у водорослей, где ни о какой функции всасывания нет и речи!).
Проясним термин “двудомные”, указанный вначале. Он означает, что женские (архегонии) и мужские (антеридии) половые органы развиваются на разных растениях. На женском гаметофите расположен архегоний, имеющий бутыльчатую форму, на мужском гаметофите развиваются антеридии. Антеридии окружают характерные крупные листья желто-бурого цвета, по которым всегда можно отличить мужской гаметофит от женского: на женском подобные листья отсутствуют.
Спорофит (бесполое поколение) у мхов располагается прямо на гаметофите, представлен коробочкой на ноже, в которой образуются споры. Коробочка способствует распространению спор на большее расстояние. Спора – возникающая в результате митоза или мейоза (в этом случае – мейоспора) клетка грибов и растений, служащая для размножения и распространения растений и грибов, способная развиться во взрослую особь.
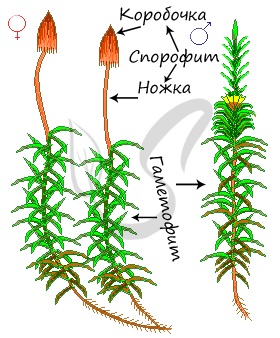
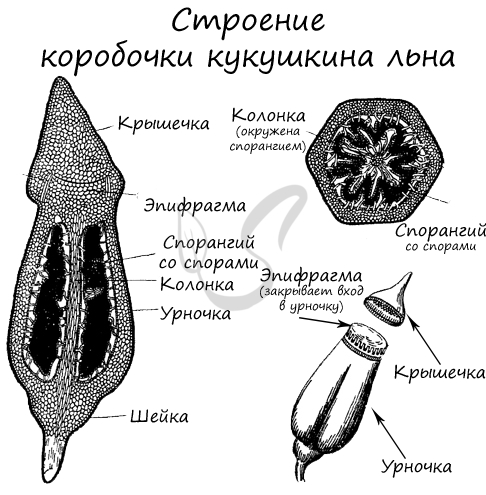
Стебелек вместе с коробочкой называется спорогон. Коробочка состоит из урночки и крышечки, сверху прикрыта колпачком. Внутри урночки находится центральный стержень – колонка, к которой крепится спорангий со спорами. Кверху колонка расширяется и образует эпифрагму, затягивающее устье коробочки. Коробочка имеет перистом – специальное отверстие для рассеивания спор.
В сухую погоду, когда споры созревают, колпачок, а за ним и крышечка, отпадают. Тонкий стебелек спорогона раскачивается от ветра, споры высыпаются.
Жизненный цикл мхов
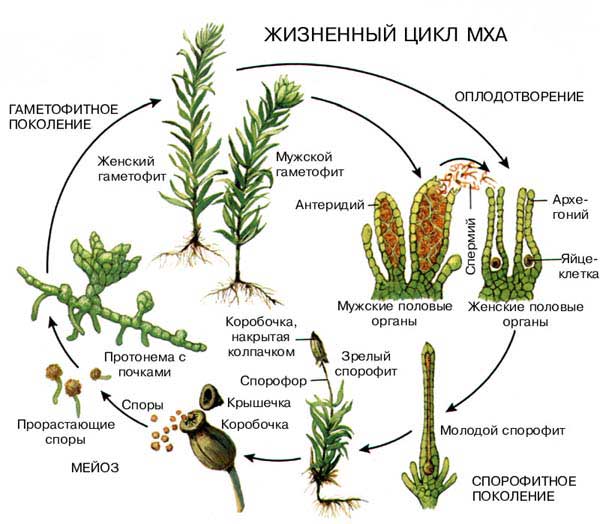
При созревании яйцеклетки (n) в архегонии (n) образуется узкий слизистый канал, по которому сперматозоид (n) может достичь яйцеклетки, которая неподвижна. Сперматозоиды образуются в другом месте – антеридиях, на мужском гаметофите, (n), и, чтобы добраться до архегониев, им обязательно нужна вода. Именно поэтому мхи обитают во влажной среде, где условия для размножения наиболее благоприятные.
Сперматозоид обладает хемотаксисом к слизи в канале архегония. Хемотаксис – движение организма под влиянием химических веществ.
В период обильных дождей сперматозоид достигает яйцеклетки в архегонии (на женском гаметофите), гаметы сливаются, и образуется зигота (2n). В дальнейшем из зиготы прорастает спорофит, имеющий вид коробочки на ножке. Отметим, что спорофит, по сути “паразитирует” на гаметофите (n), от которого зависит полностью. В коробочке спорофита из материнских клеток (2n) образуются споры (n) путем мейоза. Высыпаясь из коробочки, спора попадает в почву, где прорастает в протонему. Протонема – нитчатое образование, ранняя стадия развития гаметофита – листостебельного растения (n).
На женских гаметофитах развиваются архегонии, на мужских – антеридии, в которых образуются гаметы. Цикл замыкается. Хотелось бы еще один раз подчеркнуть: спорофит у мхов редуцирован, низведен до уровня полностью зависимого придатка гаметофита, растущего прямо на гаметофите и питающегося за счет него. Гаметофит доминирует в жизненном цикле и представлен зеленым листостебельным растением.
Теперь попробуйте самостоятельно назвать все стадии жизненного цикла мха на картинке ниже, почувствуйте себя истинным бриологом 😉
Значение мхов
В целом хозяйственное значение мхов для человека небольшое. Они не пригодны в качестве корма для домашних животных. Мох сфагнум с давних пор применяется как перевязочный материал, военные врачи применяли его в экстренных случаях, когда заканчивались вата, йод. Сфагнум обладает антисептическими свойствами: уничтожает микроорганизмы или задерживает их размножение, а также выраженной гигроскопичностью – способностью поглощать, впитывать большие количества воды, крови и других жидкостей.
Разрастание мхов вызывает переувлажнение почвы и впоследствии – заболачивание территории. Особо отметим роль мхов, как торфообразователей. Торф – горючее полезное ископаемое, образованное не полностью разложившимися остатками растений в условии болот. Его используют для производства теплоизолирующих материалов, применяют в качестве топлива. И, конечно же, мхи – звено в цепи питания (продуценты – производители органических веществ).

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Протонема, или предросток,- наиболее характерная фаза развития мхов, отличающая их от всех высших растений. Протонема вообще представляет собой преимущественно нитчатую структуру, образующуюся при прорастании споры и предшествующую развитию гаметофора в виде листостебельного побега или слоевища. У печеночников протонема представляет собой обычно короткую нить в одну или несколько клеток, из которых апикальная (верхушечная) вскоре делится продольной перегородкой на две клетки, дающие уже начало пластинчатому или листостебельпому побегу – гаметофору.
Наиболее изучена протонема мха фунарии (Funaria hygrometrica). Спора фунарии при благоприятных условиях прорастает очень скоро, и трубка, образованная зндоспорием, превращается в фототропически отрицательную нить. Эта нить со временем превращается в ризоид. Вслед за ризоидом образуется фототропически положительная разветвленная нить, нарастающая делением верхушечной клетки. Это зеленая протонема – хлоронема. Хлоронем может быть несколько; они распространяются во все стороны от споры радиально и образуют протонематическую дерновину. Апикальная клетка – единственная в нити растущая и делящаяся, так как интеркалярные клетки не растут, но могут образовывать боковые ветви.
Ветвление нити протонемы фупарии начинается только с третьей или четвертой клетки, начиная от верхушки. Ни апикальная, ни субапикальная клетка не дает ветвей, но с ростом протонемы образование ветвей продвигается вперед все в той же последовательности. Порядок ветвления зависит от наличия в нити апикального доминирования, т. е. регуляторного влияния апикальной клетки. В этом можно убедиться экспериментально, умертвив верхушечную клетку. После ее устранения порядок ветвления меняется на обратный, субапикальная клетка сразу же образует ветвь, а за ней последовательно и другие ветви. Верхняя ветвь вскоре изгибается и принимает направление роста нити, а ее верхушечная клетка занимает доминирующее влияние в регуляции ветвления нити.
Как показали опыты, характер ветвления нити зависит от того, что в апикальной клетке синтезируются гормоны – регуляторы роста, мигрирующие в нижеследующие клетки, задерживая ветвление. При взаимодействии с веществами, синтезируемыми в клетке на свету, доминирующее влияние угасает в четвертой клетке.
Протонема фунарии неоднородна. Хлоронема – это начальная стадия развития протонемы, а также боковых нитей каулонемы. Хлоронема – также начальная стадия вторичной регенеративной протонемы. Завершающая стадия развития протонемы – каулонема, на ней образуются почки гаметофоров и ризоиды. Значение ризоидов заключается в прикреплении нити протонемы к субстрату, а также в снабжении водой и минеральными солями и передаче их протонематической дерновинке. Особенно важную роль ризоиды играют у тропических моховидных – эпифиллов, растущих на листьях вечнозеленых растений. Названные фазы развития у мхов не всегда морфологически различимы и легко взаимообратимы. Каулонема, хорошо отличимая у фунарии по темноокрашенным стенкам клеток, косым поперечным перегородкам и крупным ядрам, выражена не у всех моховидных.
Нитчатая протонема известна почти у всех мхов, а также у некоторых печеночников. Внешне она похожа на нитчатую зеленую водоросль, что послужило поводом для предположения о происхождении моховидных непосредственно от зеленых водорослей. Но для этого заключения нет никаких фактических оснований. В частности, стенки клеток протонемы биохимически кардинально отличны от клеточных стенок зеленых водорослей. Таким образом, протонема у моховидных как юношеская фаза развития гаметофита – явление вполне оригинальное. Протонема обнаруживает как регрессивную, так и прогрессивную тенденцию развития. Регрессивное развитие заключается в том, что спора начинает прорастать уже в коробочке, образуя многоклеточную спору, которая или прорастает в нить (ортотрихум, дреммондия, фонтипалис), или краевая клетка многоклеточной споры развивается прямо в листостебельный побег (дикнемон).

С другой стороны, наблюдается увеличение значения протонемы в жизненном цикле мха, и протонема, в конце концов, охватывает весь цикл, за исключением продукции спорогона. Известны многие примеры так называемых протонематических мхов с постоянной протонемой (рис. 22). Из них самой знаменитой является схистостега перистая (Schistostega permata) с ее светящейся протонемой. Именно этот мох имел в виду великий немецкий поэт Гёте, философ и ботаник, вложив в уста Мефистофеля во время его прогулки с Фаустом в горах Гарца слова (перевод А. С. Лазаренко):
Не освещает ли для пира
здесь Маммон пышно свой чертог?
Гёте хорошо знал самосветящийся мох: в горах Гарца протонема его светится в полумраке пещер и расщелин в скалах. Мох изобилует в горах Средней Европы. Он и дал повод для легенд и сказок об охраняемых гномами зачарованных сокровищах, не дающихся в руки человека, так как на дневном свету свечение протонемы гаснет.
Свечение обусловлено не фосфоресценцией, а особым строением пластинчатых структур протонемы, сидящих на прямостоячих ветвях, растущих в направлении света. Пластинка, вызывающая свечение, направлена против падающего света и слагается линзовидными клетками с выпуклой передней и воронковидной задней стенкой. Именно в заднем углу клетки размещены 4-6 хлоропластов. Луч света преломляется сферической передней стенкой, направляется на хлоропласты, пройдя насквозь хлоропласт, отражается задней стенкой; вновь преломленный в передней стенке, выходит параллельно входному лучу уже как зеленый свет. При изменении угла падения световых лучей изменяется и положение хлоропластов, собирающихся снова в фокусе преломленных лучей. Фокусированием световых лучей на хлоропластах достигается оптимальный для фотосинтеза эффект в условиях рассеянного света.
Самосветящийся, или самосветный, мох широко распространен в северном полушарии. В нашей стране его можно встретить в горах Дальнего Востока и в Сибири, а на западе – от Карелии до Днепровских порогов на юге. На Украине его особенно много в Карпатах, и прекрасным зрелищем пышно осветленных чертогов Маммона можно любоваться в пещерах и скалах горы Пожижевской вблизи биологического стационара Института ботаники АН УССР.
Подобное изумрудное освещение пещер и скал в тропиках Старого и Нового Света вызывают слоевища печеночников из рода циатодиум (Cyathodium), особенно циатодиума пещерного. У него свечение обязано также отражению световых лучей линзообразными клетками верхнего слоя слоевища, такой же формы, как и в пластинке протонемы схистостеги. Самосвечение следует рассматривать как приспособление для повышения энергии фотосинтеза с помощью фокусировки рассеянных лучей света на хлорофилловые зерна.
Удивительный пример эволюции нитчатой протонемы представляет мох буксбаумия (Buxbaumia). У него протонема является его главной жизненной формой. На женской протонеме гаметофоры редуцированы и представлены коротким клеточным телом с бесцветными листьями и архегониями, а все питание спорофита обеспечивает нитчатая протонема. Мужская протонема устроена еще проще, не образует гаметофоров, а шаровидный антеридий сидит просто на ветви протонемы, прикрытый раковинообразной пластинкой из клеток протонемы же. Это наиболее далеко зашедший случай редукции гаметофора у мхов.
У мхов постоянная протонема, заменяющая облиственный побег в питании растения, особенно его спорогона, встречается довольно часто, , главным образом у клейстокарпных форм в родах эфемерум, эфемерелла, спорледера, акаулон и даже у политриховых (погонатум игольчатый). У печеночников также известна постоянная протонема, например у протоцефалозии с нитчатой и у метцгериопсиса с ветвящейся пластинчатой протонемой.
У таких мхов, как тетрафис и тетрадонциум, на нитчатой протонеме образуются клеточные пластинки, а у дифисциума – воронковидные образования, увеличивающие фотосинтезирующую поверхность протонемы. Почки гаметофоров в этих случаях образуются по краю таких пластинок. Следует добавить, что на нитчатой протонеме очень часто образуются выводковые тела, служащие для вегетативного размножения.
У андреи и сфагнума нитчатая протонема совсем не образуется, и юношеская форма гаметофита у них представлена пластинкой.
Главная роль нитчатой протонемы – это массовое образование гаметофоров, ведущее к созданию моховой дерновинки.
Жизнь растений: в 6-ти томах. – М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
Источник
МХИ, листостебельные мхи (Musci), класс отдела моховидных (Bryophyta) царства растений. Охватывает примерно 14 000 видов мелких споровых растений, встречающихся по всему миру в разнообразных средах, включая пустыни и водоемы, хотя большинство приурочено к влажным наземным местообитаниям.
Строение.
Многие мхи образуют прямостоячие неветвистые побеги, собранные в плотные дерновины или подушки. Другие, сильно ветвясь, стелются по почве или другим субстратам, например коре деревьев или скалам. Высота побегов обычно не превышает 5 см. Листорасположение спиральное; корней нет, а их функцию выполняют ризоиды.
«Стебель».
Настоящие стебли и листья у мхов отсутствуют, и соответствующие им структуры обозначают особыми терминами – каулидии и филлидии (филлоиды). У многих мхов клетки каулидия недифференцированны, у других наблюдается центральный тяж из толстостенных клеток. Они, несомненно, выполняют опорную функцию, но не эквивалентны проводящей системе – специализированным тканям, служащим для транспорта воды и питательных веществ у сосудистых растений. Большая часть воды и солей, необходимых для жизнедеятельности мхов, втягивается в их тело из внешней среды капиллярными силами через щели между филлидиями и каулидием.
«Листья».
Филлидии бывают разными по форме и размеру. Обычно они состоят всего из одного слоя клеток, но у некоторых видов по краям филлидия этих слоев несколько. Если присутствует средняя жилка толщиной в несколько клеток, она бывает одиночной, доходящей до вершины филлидия, или двойной и короткой. У некоторых видов на ней образуются пластинчатые или столбчатые выросты. Форма филлидия бывает округлой, овальной, ланцетной, продолговатой или линейной, а его край может быть цельным или зубчатым, плоским или свернутым. Эти признаки достаточно видоспецифичны и используются в таксономии.
Ризоиды.
Роль корней играют многоклеточные ветвящиеся нити – ризоиды. У молодых мхов они всасывают из почвы воду с растворенными в ней минеральными веществами, но со временем эту способность утрачивают и служат просто для закрепления растения в субстрате.
Жизненный цикл.
Зеленое фотосинтезирующее растение у мхов – это половое поколение, называемое гаметофитом. Гаметы, т.е. половые клетки, образуются на нем в особых половых органах (гаметангиях). Мужской гаметангий называется антеридием, женский – архегонием. Из оплодотворенной яйцеклетки (зиготы) развивается споровое поколение – спорофит. У мхов он практически лишен хлорофилла, остается прикрепленным к гаметофиту и получает от него питание. У спорофита каждая клетка содержит двойной (диплоидный) набор хромосом, а у гаметофита – одинарный (гаплоидный), как в гаметах. При слиянии спермия с яйцеклеткой из двух гаплоидных наборов образуется один диплоидный, необходимый для развития спорофита. У последнего при формировании спор происходит т.н. редукционное деление клеток (мейоз), каждая спора снова становится гаплоидной и может прорасти в такой же гаплоидный гаметофит.
Гаметофит.
Когда спора попадает во влажное место, она развивается сначала в ветвистую многоклеточную нить – протонему, или проросток. Оставшиеся на поверхности веточки протонемы становятся зелеными и фотосинтезирующими, а проникшие в грунт – бесцветными ризоидами. На зеленых частях проростка формируются боковые почки, из которых развиваются листостебельные побеги. Одна спора может дать целую колонию гаметофитов. У некоторых видов проростки долгоживущие, покрывающие иногда несколько квадратных дециметров грунта, у других – мелкие, исчезающие после появления облиственных побегов.
Гаметангии образуются терминально, т.е. на верхушках главного или боковых побегов. Антеридии и оогонии находятся либо на одной и той же веточке, либо на разных (иногда – даже на разных растениях) и окружены стерильными нитями – парафизами. Антеридий – это сферический или цилиндрический многоклеточный мешок, внутренние клетки которого дают по два подвижных двужгутиковых спермия. Архегоний – многоклеточная колбовидная структура. В ее основании (брюшке) находится единственная яйцеклетка, а «горлышко» (шейка) заполнено т.н. канальцевыми клетками, которые в процессе полового созревания разрушаются, превращаясь в привлекающее спермии вещество. Чтобы те попали в архегонии и произошло оплодотворение, необходима капельно-жидкая влага, например дождь или роса. Антеридий лопается, высвобождая спермии. Они плывут к шейке архегония, проникают в ее канал, и один из них сливается с яйцеклеткой, образуя диплоидную зиготу.
Спорофит.
Зигота начинает делиться еще в архегонии, который некоторое время растет вместе с формирующимся спорофитом. Когда он становится видимым невооруженным глазом, то состоит из трех частей: стопы, погруженной в брюшко архегония, тонкой ножки – спорофора и коробочки, где созревают споры. Растущий спорофит разрывает архегоний по кругу и выносит его верхнюю часть вверх в виде покрывающего коробочку колпачка (калиптры). Типичная зрелая коробочка – сложная структура, состоящая из урночки, крышечки и соединяющего их слоя специализированных толстостенных клеток – колечка. Набухая от воды, колечко отделяется от соседних частей коробочки и крышечка опадает, открывая устье урночки, которое бывает гладким или окруженным перистомом (околоустьем) из одного или двух концентрических рядов зубчиков. Эти зубцы либо плоские, либо несут от 4 до 64 поперечных гигроскопичных утолщений. Их число и форма относятся к важным таксономическим признакам мхов.
В зрелой коробочке находится множество свободных спор. Они выдуваются или вытряхиваются оттуда, разносятся ветром, водой или животными и, попав в благоприятные условия, прорастают.
Чередование поколений и эволюция.
Чередование в жизненном цикле диплоидного спорофита и гаплоидного гаметофита называется сменой поколений. Оно наблюдается у всех растений, однако, если у мхов гаметофит – хорошо заметная зеленая особь, у представителей всех прочих отделов этого царства он редуцирован до миниатюрного короткоживущего «заростка», иногда даже не способного к фотосинтезу, или вообще до группы клеток внутри спорофита. Таким образом, мхи – весьма специализированная ветвь эволюции, связанная происхождением с какими-то водорослями и, скорее всего, не давшая начало ни одной группе «высших», т.е. сосудистых, растений.
См. также СИСТЕМАТИКА РАСТЕНИЙ.
Источник