Жизненный цикл растений с чередованием у высших растений

Жизненный цикл растения состоит из трёх последовательных этапов:
- зарождение;
- развитие;
- размножение.
Он может быть простым и сложным. В качестве примера простого цикла можно привести хлореллу, которая размножается спорами. Развиваясь, эта зелёная водоросль становится вместилищем 4–8 автоспор, которые растут внутри материнского организма и покрываются собственной оболочкой. Но среди растений чаще встречается сложный цикл развития, который состоит из 2–3 простых.
Особенности жизненных циклов растений
Важным свойством всего живого является способность размножаться. Способ размножения бывает:
- половой (гаметами);
- бесполый (спорами);
- вегетативный (частью организма).
В сложных циклах при половом размножении всегда есть несколько обособленных фаз гаметы и зиготы. Гамета — это созревшая половая клетка с гаплоидным (ординарным) набором хромосом. Зигота с диплоидным (двойным) набором образуется в результате объединения двух гамет. Из зиготы развивается спорофит, который производит гаплоидные споры. Из спор — гаметофит, который бывает мужским и женским.
Для примера можно взять равноспоровый папоротник, у которого бывает две формы особей — сам папоротник (спорофит) и его заросток (гаметофит). Заросток — это потомство взрослых особей папоротника. Он существует очень короткий период, но успевает произвести на свет единственную крупнолиственную особь. Жизненный цикл растенияиз-за этой особенности размножения состоит из чередования поколений: от взрослого папоротника к заростку и снова к взрослому папоротнику.

Способы размножения
Половым способом размножается большинство растений. При этом происходит образование нового организма из зиготы после оплодотворения и объединения гамет (сингамии). Партеногенез — размножение без оплодотворения — также относится к половому способу: дочерний организм образуется из изогаметы, что роднит изогаметы и споры. Половое размножение практически всегда сочетается с другими способами — вегетативным или бесполым, так как само оно отличается низкой продуктивностью.
Одновременно этот способ и бесполое размножение встречается у папоротников, а в связке с вегетативным вариантом — у некоторых водорослей. У семенных растений формирование половой клетки происходит из одной дочерней зиготы, в результате чего этот процесс больше напоминает не размножение, а воспроизведение.
При бесполом размножении образуются зооспоры — клетки без клеточной стенки, которые у многоклеточных растений находятся в специальных спорангиях, и неподвижные клетки — апланоспоры. Самостоятельно такой способ размножения встречается в природе очень редко. Обычно он сочетается с половым или вегетативным.

Существует 2 типа спор: митоспоры, которые возникают при бесполом способе размножения, а мейоспоры — возникающие при половом. Митоспоры появляются при помощи митоза, в результате чего получается особь, похожая на материнскую. Мейоспоры образуются посредством мейоза при прорастании зиготы или в спорангиях. Для большинства растений характерны оба способа размножения, благодаря которым получаются две разные формы особей.
Вегетативный способ размножения
При вегетативном варианте размножения происходит разделение на акинеты — толстостенные клетки. Он заключается в отделении от маточника какой-то его части — выводковой почки или тельца. Таким способом размножаются некоторые низшие растения, в том числе саргассовые, бурые и красные водоросли. Вегетативно размножаются даже цветковые растения, например ряска. У некоторых из них образуются выводковые почки, которые опадают на землю и там укореняются. Также почки могут ответвляться и отделяться от материнского растения. У покрытосеменной группы растений очень часто встречается развитие побегов под землёй от корневища.
Распространение растений
Одним из финальных этапов размножения является распространение растений. В природе может быть 3 варианта расселения: зародышами, спорами и семенами. В крайне редких случаях распространение может происходить при помощи зигот. Ещё К. Линней связывал семенное и споровое распространение с тайнобрачными и явнобрачными растениями. Ко второму типу относилась группа голосеменных и покрытосеменных растений, а к первому — все остальные группы, в том числе водоросли, моховые и папоротники.
Способы размножения растений прошли длинный эволюционный путь от вегетативного к бесполому и половому. Сейчас разделение растений на споровые и семенные связывают не с распространением, а с размножением. Семенной способ выделяется в отдельную группу, так как он считается соединением размножения спорами и гаметами. Семенное размножение включает в себя несколько этапов: образование зигот, гамет, спор, зародышей и семени, а также расселение растения.
Чередование поколений
Жизнь растений в форме двух разных поколений может носить различные названия: смена форм развития, чередование поколений и т. д. Сменяемость большого папоротника и заростка в случае с равноспоровым папоротником — это пример чередования поколений, отмеченный фазами взрослого состояния форм особей. Эти две формы настолько отличаются внешне, что в них трудно узнать одно и то же растение. Заросток папоротника очень сложно заметить невооружённым глазом. У покрытосеменных растений аналог заростка — зародышевый мешок, который крайне мал и скрыт в глубине цветка. Среди некоторых групп водорослей эти формы особей похожи внешне, но совершенно различаются биологическими признаками. Чередование поколений встречается практически у всех высших растений и эволюционно развитых водорослей.

Жизненные циклы высших растений
Жизненный цикл высших растений, если не считать мохообразные, характеризуется тем, что гаметофит развит слабо, а в жизненном цикле большую часть занимает спорофит. Мохообразные растения отличаются тем, что спорофит развивается внутри женского полового органа и находится в непрерывной связи с гаметофитом. В случае с листостебельными мхами он выглядит как коробочка со спорами, растущая из верхней части гаметофита.
Остальные высшие растения обладают ярко выраженными спорофитами, которые представляют собой большие и сложные многоклеточные организмы с такими органами, как листва, стебли и корневая система. Большинство растений, которые человек представляет себе, когда говорит о хвощах, папоротниках или других группах, — спорофиты.
Жизненные циклы цветковых растений
Самыми прогрессивными в плане эволюции являются цветковые растения. Жизненный цикл цветковых растений характеризуется тем, что часто зародыш способен развиваться из неоплодотворённой яйцеклетки (апомиксис). Преобладающей формой цветковых является разноспоровый спорофит, представляющий собой растение с листьями и стеблем. Мужской гаметофит представлен пыльцевым зерном, а женский — зародышевым мешком (он развивается быстрее, чем у голосеменных). Органом как полового, так и бесполого размножения является видоизменённый побег — цветок. Зачатки семян защищены стенками завязи. Жизненный цикл развития растений этой группы заканчивается после оплодотворения и образования семени, зародыш в котором имеет запас питательных веществ и не зависит от внешних факторов.

Жизненные циклы голосеменных и покрытосеменных растений
К группе голосеменных растений относятся представители хвойных деревьев и кустарников. У большинства из них видоизменённые игольчатые листья. Жизненный цикл голосеменных растений отличается тем, что микроспоры (пыльца) образуются в мелких мужских шишках (пыльниках), а мегаспоры — в женских (семязачатках). Из микроспор образуется мужской гаметофит, а из мегаспоры — женский. Жизненный цикл растения из этой группы отличается тем, что оплодотворение происходит при помощи ветра, который доставляет пыльцу к семязачаткам. После этого внутри семяпочки начинает развиваться зародыш, а из него образуется семя. Оно лежит на семенных чешуях и ничем не прикрыто. Семя даёт новый спорофит, из которого вырастает новое растение.

Жизненный цикл покрытосеменных растений отличается тем, что у этой группы есть цветок, в котором образуются споры и происходит оплодотворение гаметофитов и развитие семян. Особенность этой группы – в защите семян, которые скрыты внутри плода и защищены от воздействий внешней среды.
Жизненный цикл споровых растений
Споровые растения не цветут, потому их также называют нецветковыми. Они бывают двух категорий:
- высшие (папоротники, хвощи, мхи, плауны);
- низшие (водоросли, лишайники).
Жизненные циклы споровых растений в зависимости от вида могут идти половым или бесполым вариантом. Они не способны размножаться половым способом без участия водной среды. Для полового размножения используется гаметофит, а для бесполого — спорофит. Существует две подгруппы споровых растений: гаплоидная и диплоидная. В гаплоидную подгруппу входят мхи, хвощи и папоротники, у которых более развит гаметофит, а спорофит формируется в виде заростка. Гаплоидная подгруппа отличается тем, что спорофит имеет в ней подчинённый статус.

Жизненные циклы растений: схемы
Мхи — представители примитивного вида высших растений. У них очень условное деление организма на стебель и листья, вместо корней — нитеобразные ризоиды. Произрастают они в болотистых, влажных местах и очень сильно испаряют влагу. Размножаются половым путём, спорофит зависит от гаметофита, споры образуются в специальной коробочке, которая находится над гаметофитом и связана с ним.
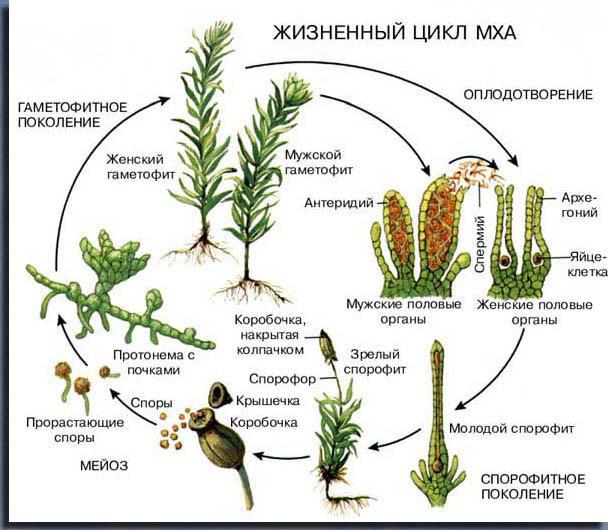
Представители папоротников обладают большими перистыми листьями (спорангии расположены на нижней стороне). У растения есть ярко выраженная корневая система, а лист на самом деле является системой ветвей, который называется вайей или предпобегом. Жизненный цикл растения группы папоротников состоит из двух фаз: половой и бесполой.
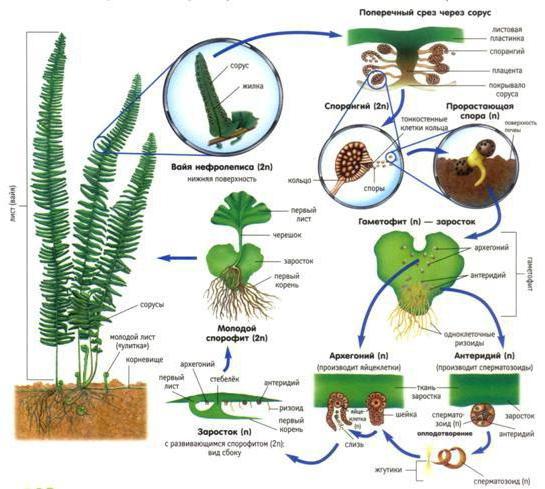
Половая фаза происходит при участии гамет, а бесполая — спор. Бесполое поколение начинается с диплоидной зиготы, а половое — с гаплоидной споры. Сменяемость этих фаз и составляет большую часть цикла.
Источник
Вернуться к теме “Высшие растения – характеристика и жизненный цикл”
Подробный конспект
Высшие растения — это многоклеточные фототрофные организмы, приспособленные к жизни в наземной среде и характеризующиеся правильным чередованием полового и бесполого поколений и наличием дифференцированных тканей и органов.
Основные признаки, отличающие высшие растения от низших:
– приспособленность к обитанию в наземной среде;
– наличие четко дифференцированных тканей, выполняющих конкретные специализированные функции;
– наличие многоклеточных органов размножения — полового (гаметангий) и бесполого (спорангий). Мужские гаметангии высших растений называются антеридиями, женские — архегониями. Гаметангии высших растений (в отличие от низших) защищены оболочками из стерильных (бесплодных) клеток и (у отдельных групп растений) могут быть редуцированы, т.е. уменьшены и упрощены;
– превращение зиготы в типичный многоклеточный зародыш, клетки которого первоначально не дифференцированы, но генетически детерминированы на специализацию в определенном направлении;
– правильное чередование двух поколений — гаплоидного полового (гаметофит), развивающегося из споры, и диплоидного бесполого (спорофит), развивающегося из зиготы;
– доминирование в жизненном цикле спорофита (у всех отделов кроме Моховидных);
– разделение тела спорофита (у большинства отделов высших растений) на специализированные вегетативные органы — корень, стебель и листья.
Всем высшим растением свойственно наличие двух жизненных фаз, закономерно сменяющих друг друга – гаметофита и спорофита. Вместе они составляют жизненный цикл высшего растения.
Гаметофит развивается из споры и представляет собой поколение, способное размножаться половым путем, т.е. слиянием двух гаплоидных клеток – гамет с образованием диплоидной зиготы. Поскольку гаметы у высших растений всегда образуются в результате митоза само тело гаметофита построено тоже из гаплоидных клеток.
Спорофит развивается из зиготы. Он размножается бесполым путем (спорами), и его тело образовано клетками с диплоидным набором хромосом. Спорофит не образует половые органы, и споры развиваются в особых многоклеточных структурах – спорангиях. Несмотря на то, что спорофит образован диплоидными клетками, споры гаплоидны т.к. образуются в процессе мейоза из материнских клеток – предшественников.

Органы растений, выполняющие функцию полового размножения.
У различных представителей растительного царства они сильно отличаются по уровню дифференциации и сложности строения. У некоторых водорослей и лишайников генеративные органы не дифференцированы на мужские и женские (морфологически не отличимы) и различаются только физиологически. В других случаях у низших растений генеративные органы подразделяются на антеридии (образующие мужские гаметы) и оогонии (образующие женские гаметы), как правило, одноклеточные. От них в процессе эволюции произошли многоклеточные генеративные органы высших растений – антеридии и архегонии. У высших споровых растений (мхов, папоротников, хвощей и плаунов) антеридии представляют собой небольшие овальные или шаровидные тельца, одетые снаружи бесплодными клетками, в. которых образуются подвижные сперматозоиды (мужские гаметы) (см. Мхи, Папоротниковидные, Плауновидные, Хвощевидные). Архегонии выглядят как небольшие бутылкообразные или колбообразные тельца, состоящие из брюшка и шейки. В брюшке помещается неподвижная яйцеклетка – женская гамета. В процессе эволюции высших растений архегонии и антеридии претерпевают упрощение (редукцию). У наиболее высокоорганизованных покрытосеменных растений, а также у некоторых голосеменных растений архегонии отсутствуют. У сосновых от архегония сохраняется яйцеклетка и несколько боковых клеток, а мужской гаметофит редуцирован до трех клеток и антеридии, и как таковой не образуется. У покрытосеменных (цветковых) растений, в связи с особенностями их развития, возникают высокоспециализированные генеративные органы – мужские гаметофиты (пыльцевые зерна), состоящие из двух клеток (генеративной и вегетативной), и сильно редуцированные женские гаметофиты – зародышевые мешки. Понятие Генеративные органы, у покрытосеменных растений часто распространяют на цветки и плоды.
Просмотров: 6304
Источник
ГЛАВА 6. РАЗМНОЖЕНИЕ РАСТЕНИЙ
6.3. Понятие о жизненных циклах и чередовании поколений у растений
Понятие о чередовании поколений возникло благодаря работе польского ботаника М. Лещик-Суминского (1820—1898), установившего в цикле развития папоротников наличие двух фаз — спороносной и половой. На заростке папоротника, который, по его мнению, является промежуточным, но вполне самостоятельным образованием между спорой и молодым растением папоротника, он нашел половые органы — антеридии со сперматозоидами и архегонии с яйцеклеткой. Это было важным научным открытием. Его книга «К истории развития папоротников» вышла в Берлине в 1848 г.
В. Гофмейстер (1824—1877) — немецкий ботаник, профессор Гейдельбергского, а затем Тюбингенского университетов высоко оценил открытие М. Лещик-Суминского. Вдохновленный широкими научными перспективами, открываемыми его работой, он принялся за сравнительно-эмбриологические исследования архегониальных растений.
В 1849 г. вышла первая работа В. Гофмейстера, в которой он ввел понятие о чередовании поколений у растений. Впоследствии В. Гофмейстер сделал величайшее открытие: он пришел к выводу, что у мохообразных, папоротникообразных и голосеменных происходит правильная смена (чередование) двух поколений — полового и бесполого. Оба эти поколения отличаются друг от друга формой, способом питания, биологическими и морфологическими особенностями. Свои представления об индивидуальном развитии изученных таксонов он изложил в капитальном труде «Сравнительное исследование над прорастанием, развитием и плодообразованием высших споровых растений и образованием семян у хвойных», опубликованном в 1851 г. в Лейпциге.
В ряде работ, вышедших в начале ХХ в. (1911, 1927), профессор Московского университета Л. И. Курсанов описал чередование поколений у зеленых и бурых водорослей.
Итак, жизненный цикл — это свойственная различным таксонам последовательность развития от какого-нибудь этапа до его повторения. Например, у семенных растений жизненный цикл продолжается от семени до семени; у плаунообразных и других споровых растений — от споры до споры. В жизненном цикле высших растений может доминировать или гаметофит (только у мохообразных), или спорофит (у всех высших споровых растений, а также у голосеменных и покрытосеменных).
Схема жизненного цикла, характерная для равноспоровых папоротников, представлена на рис. 138. Из схемы видно, что в жизненном цикле процесс спорообразования сменяется половым процессом, и как следствие этого, бесполое поколение (спорофит) сменяется половым (гаметофитом).
Рис. 138. Схема жизненного цикла у равноспоровых папоротников (геторофазное гетероморфное чередование поколений)

У плаунообразных, хвощеобразных и папоротникообразных спорофит и гаметофит существуют независимо друг от друга, т. е. разделены пространственно и представляют собой два поколения одного и того же вида.
У мохообразных доминирует гаметофит. Спорофит развивается на гаметофите и частично питается за его счет. У голосеменных и покрытосеменных растений, для которых характерна разноспоровость, доминирует спорофит. Гаметофиты формируются на материнских организмах, развиваются за счет спорофита, сильно редуцированы и никогда не покидают материнского растения.
Таким образом, под поколением следует понимать совокупность органических форм, возникающих в результате какого-либо репродуктивного процесса и генетически связанных с предками и потомками. Явление, связанное со сменой репродуктивных процессов и приводящее к образованию в жизненном цикле форм, различающихся морфологически, цитологически и функционально, называется чередованием поколений, или сменой фаз развития.
6.3.1. Общие закономерности чередования поколений (смена фаз развития)
Морфологические особенности спорофитов и гаметофитов у представителей различных таксономических групп, их взаимная связь друг с другом, а также закономерности смены фаз развития (чередование поколений) различны.
Чередование поколений бывает факультативным и облигатным.
У примитивных водорослей чередование поколений носит факультативный характер и чаще всего связано со сменой экологических факторов (понижением температуры воды, подсыханием водоема и т. п.). Так, например, пресноводная нитчатая водоросль улотрикс (Ulothrix) может многократно формировать 4-жгутиковые зооспоры, которые, проплавав некоторое время в воде, оседают на дно, утрачивают жгутики и прорастают в новые гетероталличные нити улотрикса. Осенью в клетках (одноклеточных гаметангиях) улотрикса формируются изогаметы и в результате процесса оплодотворения образуется зигота. Она «одевается» прочной оболочкой, оседает на дно, где и сохраняется до весны.
Схематично факультативное чередование поколений у улотрикса выглядит так: n… n…. n… 2n… n… n и т. д.
Однако у подавляющего большинства водорослей и у всех без исключения высших растений в жизненном цикле наблюдается строго закономерная смена спорообразования и полового процесса и происходит облигатное (обязательное) чередование поколений. Схематично его можно представить так: n… 2n… n… 2n… n… 2n, если в жизненном цикле доминирует гаметофит; или 2n… n… 2n… n… 2n… n, если в жизненном цикле доминирует спорофит.
Переход от спорофита к гаметофиту происходит в результате редукционного деления (мейоза) спороцитов (материнских клеток спор), которое происходит в спорангиях и предшествует образованию спор. Из спор развивается гаметофит, на котором формируются половые клетки — гаметы. Переход от гаметофита к спорофиту — следствие процесса оплодотворения яйцеклетки сперматозоидом (у высших споровых растений) или спермием (у высокоорганизованных голосеменных и покрытосеменных растений), в результате чего образуется зигота, восстанавливается диплоидный набор хромосом и начинает из зиготы формироваться спорофит.
Чередование поколений может быть гомофазным и гетерофазным. Эти термины введены П. Хартманом в 1939 г.
При гомофазном чередовании поколений спорообразование и половой процесс происходят у примитивных гаплоидных водорослей при одной и той же ядерной фазе, иначе говоря, смена репродуктивных процессов не влечет за собой изменений ядерной фазы. Например, зооспоры и изогаметы у улотрикса в различные периоды формируются в результате митоза на одних и тех же гетероталличных гаплоидных нитях (рис. 139).
Рис. 139. Схема гомофазного чередования поколений у улотрикса (Ulotrix sp.)

При гетерофазном чередовании поколений смена репродуктивного процесса (или, как неточно говорят, способа размножения — именно так полагал В. Гофмейстер) всегда влечет за собой смену ядерной фазы, т. е. спорообразование происходит на спорофите и сопровождается редукцией числа хромосом,
а образование гамет осуществляется на гаметофите, и в результате процесса оплодотворения число хромосом восстанавливается (см. рис. 138).
Гетерофазное чередование поколений может быть изоморфным и гетероморфным. При изоморфном чередовании поколений гаметофит и спорофит морфологически не различимы, но они отличаются по цитологическим, физиолого-биохимическим и функциональным свойствам (рис. 140). Изоморфное чередование поколений встречается довольно редко и только у водорослей. Оно характерно для зеленых водорослей (ульва — Ulva, энтероморфа — Еnteromorpha, кладофора — Cladophora), а также у представителей ряда порядков бурых и большинства красных водорослей.
Рис. 140. Схема гетерофазного изоморфного чередования поколений у ульвы (Ulva sp.)

У высокоорганизованных водорослей (бурых, некоторых зеленых и красных) и всех без исключения высших растений чередование поколений гетерофазное гетероморфное, т. е. у большинства растительных организмов гаметофит и спорофит, кроме цитологических, физиолого-биохимических и функциональных различий, имеют четкие морфологические различия (см. рис. 138).
Соотношение полового и бесполого поколений у различных водорослей и высших растений различно. В зависимости от уровня развития гаметофита и спорофита чередование поколений может быть гаплогомофазным, диплогомофазным, гаплодиплофазным и диплогаплофазным.
У видов с гаплогомофазным циклом развития доминирует гаметофит, диплоидна только зигота (примитивные зеленые водоросли). Для таких особей характерен начальный, или зиготный, тип мейоза, т. е. редукционно делится только ядро зиготы, с последующим цитокинезом и образованием апланоспор.
При диплогомофазном чередовании поколений доминирует спорофит, гаплоидны только гаметы (диатомовые — Bacillariophyta, из бурых — представители порядка фукусовые — Fucales, зеленые водоросли, имеющие сифональное строение). В этом случае мейоз предшествует образованию гамет. Такой тип мейоза называется конечным, или гаметным. Он характерен для животных и крайне редок у растений.
При гаплодиплофазном чередовании поколений доминирует гаметофит. У водорослей спорофит существует самостоятельно и может морфологически не отличаться от гаметофита (Ulva), или имеет микроскопическое строение (акросифония из Chlorophyta), или спорофит развивается на гаметофите (филлофора Броди из Rhodophyta). У высших растений только мохообразные характеризуются гаплодиплофазным циклом развития (рис. 141). При этом спорофит в виде коробочки всегда развивается на гаметофите, частично питается за его счет, хотя на ранних этапах развития имеет зеленую окраску и, следовательно, способен к фотосинтезу. Мохообразные являются равноспоровыми растениями, однако гаметофиты их (т. е. сами растения) могут быть как однодомными, так и двудомными. У однодомных растений антеридии и архегони образуются на одном и том же растении (Sphagnum squarrosum, S. compactum, Mnium cuspidatum). У двудомных растений половые органы одного типа (антеридии или архегонии) образуются на разных особях (Marchantía polymorpha, Sphagnum magellaniсum, Polytrichum commune).
Рис. 141. Схема гаплодиплофазного чередования поколений у мохообразных
При диплогаплофазном типе развития в жизненном цикле высших растений доминирует спорофит. Гаметофиты могут существовать независимо от спорофитов (плаунообразные, хвощеобразные, папоротникообразные), или они формируются на спорофитах (голосеменные, покрытосеменные).
При гаплодиплофазном и диплогаплофазном жизненных циклах мейоз предшествует образованию спор. Такой тип мейоза называется споровым, или промежуточным. Он характерен для всех без исключения высших растений, а также встречается у многих водорослей.
Растения с гаплодиплофазным и диплогаплофазным циклами развития могут быть как равноспоровыми, так и разноспоровыми.
У разноспоровых (селягинелловые, сальвиниевые) высших растений по сравнению с равноспоровыми, заростки сильно редуцированы (рис. 142). Однако это явление имеет большое биологическое значение.
Рис. 142. Спорофит, спорангии, споры, гаметофиты селягинеллы (Selagmella sp.): А—внешний вид спорофита; Б—микроспорангий с микроспорами; В—микроспора; Г— мужской гаметофит; Д—макроспорангий с макроспорами; Е—макроспора; Ж—женский гаметофит: 1—оболочка микроспоры; 2—оболочка макроспоры

Заростки у селягинелловых, сальвиниевых развиваются почти полностью внутри оболочки споры, следовательно, они надежно защищены от внешней среды (от неблагоприятных условий) и быстро формируются в процессе онтогенеза, что очень важно с эволюционной точки зрения.
У семенных растений (для них характерна гетероспория) при крайней степени редукции мужского гаметофита (пыльцевых зерен, пыльцы) расширяются его функции. Наряду с типичной функцией образования гамет (половой функцией) мужской гаметофит выполняет транспортную функцию. Мужские гаметы доставляются к яйцеклетке (в архегоний у голосеменных растений или в зародышевый мешок у покрытосеменных) пыльцевой трубкой, которая формируется у мужского гаметофита.
В биологии развития разноспоровых растений особого внимания заслуживают три особенности:
1) дифференциация спор — физиологическая и морфологическая, причем функцию расселения вида у высших разноспоровых осуществляет мегаспора;
2) надежная защита гаметофитов от неблагоприятных условий;
3) редукция гаметофитов, и как следствие этого, их быстрое развитие в онтогенезе.
В процессе эволюции у семенных растений мегаспора утрачивает функцию расселения, оно осуществляется семенами. У них более выражены черты приспособления к условиям существования на суше: происходит крайняя редукция гаметофитов, особенно мужских; наряду со сперматозоидами (саговник — Cycas, гинкго — Ginкgo) появляются спермии (хвойные, все покрытосеменные); при прорастании мужского гаметофита образуется пыльцевая трубка и возникает сифоногамия, которая делает процесс оплодотворения не зависящим от наличия воды.
Конечный результат появления разноспоровости — сокращение продолжительности жизненного цикла растительного организма.
6.3.2. Представление об эволюции жизненных циклов растений
У различных таксонов наблюдаются некоторые специфические особенности жизненного цикла, но в целом для растений основные направления эволюции жизненных циклов вырисовываются довольно четко.
1. На ранних этапах эволюции все особи выполняли и вегетативные, и репродуктивные функции. Половой и бесполый процессы равнозначны. Одна и та же особь потенциально могла формировать и споры, и гаметы (см. рис. 139).
2. Постепенно происходит дифференцировка особей по способам репродуктивных процессов на спорофиты и гаметофиты, но морфологически они не различаются (см. рис. 140).
3. Появляются морфологические различия между гаметофитом и спорофитом, но доминирует гаметофит. Спорофит развивается на гаметофите (см. рис. 141).
4. Спорофит отделяется от гаметофита, т. е. происходит преодоление зависимости спорофита от гаметофита (см. рис. 134, 135, 136).
5. В жизненном цикле закрепляется господствующее положение спорофита. Первоначально для него характерна изоспория (равноспоровость). Спора выполняет функцию расселения вида.
6. Появляется гетероспория (разноспоровость), что приводит к половой дифференцировке гаметофитов. Функцию расселения вида выполняет только макроспора (мегаспора) (см. рис. 142).
7. В связи с гетероспорией происходит редукция гаметофитов, что ускоряет процесс их развития. В то же время расширяются функции мужского гаметофита: кроме образования гамет и выполнения половой функции он приобретает транспортную функцию (доставляет мужские гаметы к яйцеклетке).
8. Постепенно устанавливается строгая последовательность смены спорообразования и полового процесса.
9. У высокоорганизованных высших растений (голосеменные и покрытосеменные) гаметофиты достигают крайней степени редукции, развиваются на спорофитах. Макроспора утрачивает функцию расселения вида, она никогда не покидает материнского растения.
10. На основании сочетания спорообразования и полового процесса возникает семя — сложное образование, в котором формируется зачаток нового организма (новый спорофит) и сохраняются остатки материнского спорофита. У покрытосеменных это семенная кожура и у некоторых растений питательная ткань — перисперм (гвоздичные, маревые). У голосеменных в семени есть еще и хорошо развитый и превращенный в трофическую ткань женский гаметофит (первичный эндосперм).
Чередование поколений имеет эволюционное и биологическое значение.
Большое эволюционное значение чередования поколений состоит в том, что смена репродуктивных процессов привела к доминированию в жизненном цикле спорофита, который чрезвычайно сложно устроен и наилучшим образом приспособлен к жизни в разнообразных условиях.
Биологическое значение чередования поколений заключается в том, что постепенно, с усложнением морфофизиологического строения спорофита и редукцией гаметофита совершенствовался половой процесс, и его конечный этап — оплодотворение, которое стало осуществляться без наличия капельножидкой среды, что очень важно в условиях наземного существования.
Источник