Жизненный цикл несовершенных грибов

Дейтеромицеты, дейтеромицеты или deuteromycotas, Также известные как несовершенные грибы, они являются грибами, которые не знают или не знают о половой фазе (отсюда и термин «несовершенный»). Этот таксон, содержащий около 25 000 видов, в настоящее время не считается действительным.
В большинстве случаев они являются сапрофитами, то есть питаются разлагающимся органическим веществом. Некоторые виды могут быть паразитирующими на растениях или животных, включая людей.
Некоторые несовершенные грибы имеют коммерческое значение. Его основное использование в процессах промышленной ферментации продуктов питания и напитков. Они также используются для производства лекарств и биологического контроля вредителей.
индекс
- 1 Характеристики
- 2 Таксономия
- 3 Хабитат
- 4 жизненных цикла
- 5 Питание
- 6 Репродукция
- 7 болезней
- 7.1 В растениях
- 7.2 У животных
- 7.3 У людей
- 8 Использование / применение
- 9 Ссылки
черты
Несовершенные грибы представляют большое разнообразие форм тела. Большинство из них похожи на бесполую фазу аскомицетов. Другие могут быть перепутаны с базидиомицетами или с зигомицетами. Некоторые виды одноклеточные.
Мицелий образован хорошо развитыми гифами, межклеточными или внутриклеточными. Гифы сильно разветвленные, многоядерные и имеют простые перегородки пор. Основным компонентом его клеточной стенки является хитин-глюкан.
Размножение бесполое, как правило, с помощью не жгутиконосых спор, называемых конидиями. Конидии могут иметь форму сферы, цилиндра, звезды, спирали и др..
Эти споры образуются в структурах, называемых конидиеносцами. Конидиеносцы могут быть простыми или разветвленными. Они могут расти в одиночку или в группах, образующих сферические плодоношения.
В некоторых случаях плодоношения имеют форму бутылок, в таких случаях они называются пикнидами. Если они принимают форму блюдца, их называют acérvulos.
таксономия
Традиционная классификация грибов основана главным образом на характеристиках плодовых тел и спор. Эти структуры производятся во время полового размножения.
Из-за этого грибы, которые не представляли или были неизвестны, этот тип размножения были включены в тип дейтеромицетов. В настоящее время насчитывается около 15 000 видов дейтеромицетов, сгруппированных в 2600 родов..
Многие авторы утверждают, что дейтеромицеты действительно являются аскомицетами, половая фаза которых неизвестна, вероятно, потому, что это происходит очень редко. Также возможно, что эта фаза была потеряна в ходе эволюционного процесса..
Несколько фактов, кажется, поддерживают эту теорию: большинство Deuteromycetes очень похоже на бесполую фазу (анаморфы) Ascomycetes; большинство дейтеромицетов, для которых была обнаружена их половая фаза (теломорфы), оказались аскомицетами, такие же результаты были обнаружены при перекрестном размножении в лаборатории и при молекулярных исследованиях.
Многие Deuteromycetes, которые были перемещены в другие таксоны, имели известную и описанную половую фазу как другой вид. В этих случаях они сохранили оба названия, в результате чего виды с двумя научными названиями.
Теломорф получает название аскомицетного «вида» (или соответствующей группы), а анаморфа – название, которое он получил как несовершенный гриб. Однако существует тенденция принимать только одно имя.
среда обитания
Дейтеромицеты – это повсеместно распространенные организмы. Хотя большинство видов встречается в почвах, некоторые предназначены для водной среды, а другие – для воздуха..
Некоторые организмы живут в различных средах, другие имеют более ограниченную среду обитания. Например, некоторые виды растут только в гниющей древесине, другие – в подстилке или в обугленной древесине..
Некоторые являются специфическими паразитами для одного вида хозяина, другие могут паразитировать на нескольких разных видах.
Жизненные циклы
Дейтеромицеты также известны как “бесполые грибы” и “конидиальные грибы”, потому что в их жизненном цикле присутствует только бесполая фаза. Остальные грибы могут размножаться как половым, так и бесполым, поэтому их жизненные циклы более сложны..
Споры, попадающие в окружающую среду, переносятся ветром, водой или каким-либо биологическим вектором и после оседания на соответствующем субстрате прорастают. Как только спора прорастает, новый гриб начинает расти и развиваться.
Если гриб растет на субстрате, он достигнет зрелости и размножится в том месте, где он прорастал. Если это эндопаразит, он должен выделять ферменты, которые позволяют ему разрушать защитное покрытие своего хозяина..
Растительные паразитарные грибы выделяют ферменты, разрушающие клеточную стенку. Те, которые паразитируют на насекомых или энтомопатогенах, выделяют хитиназы. Дерматофиты, с другой стороны, выделяют кератиназы.
Как только половая зрелость достигнута, они производят новые споры в конидиеносцах. В случае эндопаразитов, когда они созревают, они проецируют конидиеносцы вне хозяина..
После получения спор они попадают в окружающую среду, откуда они будут транспортироваться, чтобы получить место для прорастания и начала нового цикла..
питание
Большинство дейтеромицетов питаются разлагающимся органическим веществом. Другие виды паразитируют на растениях или животных.
Сапрофитные виды питаются с помощью ферментов, которые выделяют среду. Эти ферменты переваривают и растворяют органическое вещество, позволяя его адсорбции грибами.
Органическое вещество может быть растительного происхождения, как остатки листьев, стволов, обугленные остатки растений, разлагающиеся плоды. Это может также быть животного происхождения: трупы, кости, рога, кал, среди других.
Паразитические виды должны производить и выделять вещества, которые позволяют им разрушать клеточные стенки, экзоскелеты или кутикулу их хозяев, чтобы проникать в них и питаться их жизненно важными жидкостями или тканями..
воспроизведение
Дейтеромицеты размножаются бесполым путем образования спор, фрагментации и / или почкования мицелия. Споруляция является наиболее распространенной формой бесполого размножения. Споры, или конидии, бесполые и афлагелатированные и образуются в конидиеносце путем митотического деления.
Фрагментация – это самопроизвольный разрыв гифы, при котором образуются частицы гифы, которые отделяются от гриба и способны развиваться и образовывать новые организмы..
Во время зачатка путем деления клеток гифы образуется зародыш, который будет увеличиваться в размерах и развиваться, не отделяясь от гриба. Когда это развилось, это отделяется от своего родителя и формирует новое независимое тело.
Как механизм увеличения их генетической изменчивости, в редких случаях у дейтеромицетов может быть парасексуальный цикл. В этом цикле обмен генетическим материалом происходит внутри одного и того же организма..
Во время парасексуального цикла происходят следующие события: образование гетерокариотического мицелия, слияние некоторых пар гаплоидных ядер с образованием новых диплоидных ядер, митоз обоих типов ядер, скрещивание диплоидных ядер во время митоза и гаплоидизация некоторых диплоидных ядер.
Гаплоидизация – это процесс митотического деления, во время которого происходит сшивание и уменьшение количества хромосом. При этом процессе гаплоидные ядра могут быть получены из диплоидных ядер без мейоза.
болезни
В растениях
Многие виды этой группы вызывают заболевания у растений. Гниль кукурузы, томатов и хлопка, некоторые формы антракноза, язвы (язвы) и ожоги листьев – некоторые из заболеваний, приписываемых дейтеромицетам.
У животных
Некоторые виды дейтеромицетов являются энтомопатогенными, которые могут вызывать достаточно тяжелые эпизоотии, которые почти полностью уничтожают популяции насекомых..
Грибок rhizium anisopliae нападает на термитов вида Heterotermes tenuis, которые в свою очередь влияют на резину (Гевея бразильская) в колумбийской Амазонии.
Deuteromycetes рода Culicinomyces паразитировать комаров рода малярийный комар. Другие роды грибов, такие как Beauveria, rhizium и Tolypocladium они также нападают на комаров.
Дерматофитными грибами, поражающими животных, являются в основном дейтеромицеты, принадлежащие к родам. Microsporum и Trichophyton.
Функциональная классификация дерматофитов разделяет их на зоофилы, которые в основном поражают животных, но могут передаваться человеку; антропофильные, они встречаются в основном у людей, редко передаются животным; и геофилики, которые находятся в основном в почве, связаны с останками животных, которые содержат кератин, заражают как людей, так и животных.
У крупного рогатого скота дерматофитоз очень распространен в странах с холодным климатом, потому что животные содержатся в конюшне в течение длительных периодов времени. Большинство повреждений у здоровых животных заживают спонтанно в течение одного-нескольких месяцев.
У людей
Основным действием дейтеромицетов на человека является дерматофитоз. Виды Epidermophyton floccosum Он является патогенным для человека и в первую очередь ответственен за “ногу спортсмена” и tinea cruris. Другими дерматофитозами являются различные виды опоясывающего лишая (миндалины, телесные, борода, лицевые, крестные, стопы, кисти, паховые).
Большинство дерматофитозов не являются серьезными у здоровых людей, но могут быть более серьезными у людей с ослабленной иммунной системой.
В этих случаях могут возникнуть атипичные и агрессивные инфекции, обширный дерматит и подкожные абсцессы. Другая скрытая опасность заключается в том, что оппортунистические бактерии могут вызывать целлюлит в коже, поврежденной межпальцевым дерматофитозом..
Использование / приложения
Некоторые дейтеромицеты используются в промышленных целях, главным образом для ферментации продуктов питания и напитков. Их также используют для получения лекарств, например пенициллина, полученного из гриба пеницилл.
Некоторые виды используются для биологического контроля над насекомыми (энтомопатогены). Эти грибы имеют определенные преимущества перед другими микробными агентами контроля, такими как бактерии, простейшие и вирусы.
Несовершенные грибы / дейтеромицеты и другие грибы способны поражать все стадии развития насекомых. Они также могут атаковать виды насекомых, которые обычно не подвержены заражению бактериями и вирусами..
ссылки
- М. Арабацис, А. Велеграки (2013). Цикл полового размножения у оппортунистического возбудителя человека Aspergillus terreus. Mycologia.
- М. Блэквелл, Д. Хиббетт, Дж. Тейлор, Дж. Спатафора (2006). Исследовательские координационные сети: филогения для царства грибов (Deep Hypha). Mycologia.
- Грибки несовершенны. В википедии. Получено 2 сентября 2018 г. с сайта en.wikipedia.org
- М. Мора, А. Кастильо, М. Фрага (2017). Классификация и механизм заражения энтомопатогенных грибов. Arquivos do Instituto Biológico.
- J.L. Питт, J.W. Тейлор (2014). Aspergillus, Его сексуальные государства и новый международный кодекс номенклатуры. Mycologia.
- Д. Сикард, П.С. Pennings, C. Grandclément, J. Acosta, O Kaltz, J. Shykoff (2007). Специализация и локализация грибкового паразита на двух видах растений-хозяев, выявленная по двум признакам пригодности. эволюция.
- J. Guarro, J. Gene, A.M. Stchigel (1999). События в таксономии грибов. Клиническая Микробиология Отзывы.
Источник
Дейтеромице́ты (лат. Deuteromycóta, Deuteromycotína), также несоверше́нные грибы́ (лат. Fungi Imperfecti), митоспо́ровые грибы (англ. mitosporic fungi), анамо́рфные грибы (англ. anamorphic fungi) – грибы, размножающиеся посредством пропагул, образующихся без участия процессов мейотического деления. Как правило, эти пропагулы представляют собой конидии, реже образуются из недифференцированного мицелия.
Многие дейтеромицеты представляют стадии жизненного цикла совершенных аскомицетовых и базидиомицетовых грибов и при некоторых условиях переходят в стадию, способную образовывать споры в процессе мейоза. У многих дейтеромицетов совершенная форма, вероятно, существует, однако пока не определена.
В настоящее время для множества грибов устанавливается связь между дейтеромицетовыми и совершенными формами. При невозможности экспериментально подтвердить связь анаморфы и телеоморфы (совершенной формы) используются данные молекулярного анализа ДНК. Вероятно, для ряда дейтеромицетов никогда не будет обнаружена совершенная стадия, полностью выпавшая из их цикла развития. Положение таких грибов в системе, основанной на телеоморфах, может быть определено только молекулярно-генетическими методами.
Исторически дейтеромицеты описывались в ранге отдела, подотдела или класса. В первом случае в составе группы выделялись следующие классы (в настоящее время эти названия продолжают использоваться на практике для морфологического определения грибов):
- Гифомицеты (Hyphomycetes Fr., 1821) – формы, образующие конидии непосредственно из вегетативных гиф или из пучков агрегированных гиф – синнем или спородохиев, но не в специализированных плодовых телах;
- Целомицеты (Coelomycetes Grove, 1935) – формы, образующие конидии в специализированных конидиомах – пикнидиях, пикнотириях, ацервулах;
- Агономицеты (Agonomycetes Ainsw., 1966) – формы, не образующие конидий и существующие в виде вегетативного мицелия, на котором образуются хламидоспоры, склероции или подобные образования.
Дейтеромицеты – наиболее широко распространённые грибы. Многие из них имеют большую экономическую значимость. Ряд видов вызывают болезни сельскохозяйственных культур – например, роды Botrytis, Verticillium, Cladosporium, порчу пищевых продуктов – роды Fusarium, Trichothecium. Ряд представителей используются в микобиологической промышленности в качестве продуцентов антибиотиков и ферментов – Aspergillus, Penicillium.
Некоторые представители являются патогенами человека, другие выделяют различные токсины – Aspergillus, Fusarium.
Обзор подходов к классификации[править | править код]
Уже Пьер Андреа Саккардо в Sylloge fungorum описывал дейтеромицеты (в ранге класса) как искусственную гетерогенную группу, включающую бесполые стадии грибов. На протяжении длительного времени дейтеромицеты классифицировали исключительно на основании морфологических признаков. Существует множество таких систем классификации.
Впервые попытку создать классификацию грибов, в которой учитывались связи дейтеромицетов с половыми стадиями, предпринял в 1958 году Кэйсукэ Тубаки. Стали появляться классификации дейтеромицетов, учитывавшие морфологию известных телеоморф. Одной из основных проблем, с которыми столкнулись авторы таких классификаций, являлся тот факт, что к тому времени лишь для крайне незначительной доли дейтеромицетов была установлена связь с совершенными грибами. Наиболее известные из таких классификаций – системы Латтрелла и Саттона.
Э. С. Латтрелл (1979) выделил в пределах класса Deuteromycetes два подкласса – Ascodeuteromycetidae и Basidiodeuteromycetidae. В первый из них автор отнёс грибы с простыми порами септ, двуслойной клеточной стенкой, и гифами без пряжек, ко второму – грибы с долипоровыми септами, ламеллярным строением клеточной стенки и наличием пряжек на мицелии (как правило).
Б. Ч. Саттон (1980) на основании способа возникновения конидий разделил подотдел Deuteromycotina на два класса – Blastodeuteromycetes и Thellodeuteromycetes. Подклассы он выделил на основании различий в способе формирования оболочек конидий: Holoblastomycetidae и Enteroblastomycetidae в составе первого класса, Holothallomycetidae и Enterothallomycetidae в пределах второго.
Литература[править | править код]
- Дудка И. А. Проблема вида и таксономических критериев анаморф «высших» грибов // Проблемы вида и рода у грибов / под ред. Э. Пармасто. – Таллин, 1986. – С. 67-78.
- Anamorphic fungi // Dictionary of the Fungi / ed. by P. M. Kirk, P. F. Cannon, D. W. Minter & J. A. Stalpers. – 10th Edition. – 2008. – P. 28-31.
Источник
Дейтеромицеты, пли несовершенные грибы, наряду с аскомицетами и базидиомицетами представляют один из крупнейших классов грибов (в нем около 30% всех известных видов). Этот класс объединяет грибы с септированным мицелием, весь жизненный цикл которых обычно проходит в гаплоидной стадии, без смены ядерных фаз. Они размножаются только бесполым путем – конидиями, а половые (совершенные) стадии у них отсутствуют.
Дейтеромицеты существенно отличаются от других классов грибов, представители которых имеют обычно общих предков. Это заведомо гетерогенная группа, виды которой связаны но происхождению с разными группами из двух классов – аскомицетов и базидиомицетов. Некоторые микологи, подчеркивая это отличие несовершенных грибов, называют их формальным классом.
Хотя в системе несовершенных грибов мы пользуемся теми же таксономическими категориями, как и для других групп грибов, в них вкладывается иной смысл – это искусственные, формальные группы, выделяемые на основе чисто внешнего сходства оргапизмов, а не их родства. Например, род дейтеромицетов не соответствует истинному роду (группе близкородственных видов). Он представляет искусственную группу, объединяющую виды со сходными конидиальными спороношениями, называемую часто, в отличие от родов других организмов, формальным родом.
Мы знаем, что часто один род аскомицетов включает виды с разными конидиальными стадиями. Так, у рода микосферелла известны конидиальные спороношения из родов рамулярия (Ramularia), церкоспора (Cercospora), септория (Septoria) и другие, а у рода нектрия конидиальные спороношения из родов акремониум (Acremonium), фузариум (Fusarium), туберкулярия (Tubercularia), цилиндрокарпон (Суlindrocarpon).
С другой стороны, конидиальные спороношения различных родов аскомицетов, даже относящихся к разным порядкам, часто бывают очень похожи. В результате этого в один род дейтеромицетов нередко входят виды, связанные по происхождению с представителями нетолько разных родов, но и порядков и даже классов высших грибов. Род акремониум, например, связан с совершенными стадиями эмерицеллопсис (порядок эуроциевых), хетомиум (порядок сферейных), нектрия (порядок гипокрейных) и кордицепс (порядок спорыньевых), а конидиальные стадии типа эдоцефалюм (Oedocephalum) известны как у дискомицетов, так и у некоторых базидиомицетов. Связи с представителями разных классов установлены и у некоторых родов аспорогенных дрожжей, например кандида (Candida).
Вегетативное тело дейтеромицетов – хорошо развитый, ветвящийся, гаплоидный мицелий, состоящий- обычно из многоядерных клеток. В мицелии всегда присутствуют септы (перегородки), обычно с простыми порами, как у аскомицетов (рис. 51). Можно предположить, что имеются дейтеромицеты и с септами типа, характерного для базидиомицетов, но они пока не изучены методом электронной микроскопии. У аспорогенных дрожжей мицелия нет, а вегетативное тело представлено почкующимися клетками.
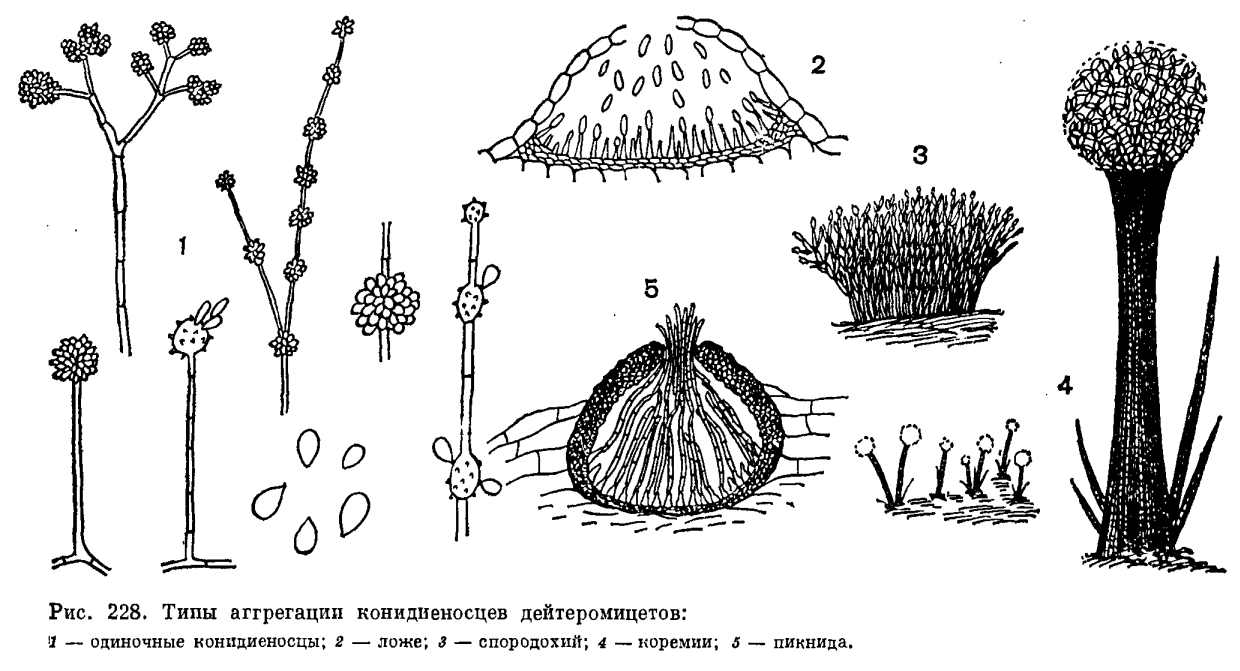
У большинства дейтеромицетов размножение происходит при помощи конидий. Лишь у немногих из них конидиальное спороношение отсутствует. Такие грибы часто образуют склероции (Sclerotium, Rhizoctonia), а иногда встречаются только в виде стерильных мицелиев. Конидии (споры бесполого размножения) образуются на гаплоидном мицелии, на многоклеточных, реже одноклеточных конидиеносцах, представляющих ветви мицелия, обычно поднимающиеся над ним. Они могут быть малодифференцированными, не отличающимися от вегетативных гиф мицелия, но чаще хорошо развиты. Увеличение продукции конидий достигается путем ветвления конидиеносцев различным образом (мутовчатое, моноподиальное, симподиальное, дихотомическое, рис. 228), образования их расширений или вздутий, несущих группы спор и расположенных на вершине конидиеносца (Aspergillus, Oedocephalum, рис. 228) или интеркалярно (Arthrys, Gonatobotrys, рис. 228), а также образованием конидий в длинных акропетальных или базипетальных цепочках (рис. 230).
, 
Многие дейтеромицеты образуют конидиеносцы, объединенные в группы на мицелии или внутри пикнид. Простейшая форма такой аггрегации конидиеносцев – коремии. Многочисленные конидиеносцы развиваются тесно сближенным пучком, обычно склеиваясь своими боками, а иногда срастаясь. В результате этого образуется компактная колонка, на вершине которой на веточках конидиеносцев синхронно развиваются конидии (рис. 228).
Коремии характерны для семейства стильбелловых (Stilbellaceae) порядка гифомицетов, а также для некоторых пенициллов. У многих дейтеромицетов конидиеносцы образуют слой на поверхности выпуклого сплетения гиф или стромы в виде подушечки; Этот тип конидиального спороношения называют спородохиями (рис. 228). Если конидиальные спороношения такого типа имеют слизистую или желеобразную консистенцию, а в основании более рыхлое сплетение гиф мицелия, их называют пиониотами. Конидии, образующиеся в пионнотах, погружены в слизь.
Ложа (ацервулы) по характеру объединения конидиеносцев напоминают спородохии, но конидиеносцы в них образуют тесный слой не на выпуклой строме, а на более или менее плоском сплетении гиф. Ложа часто встречаются у дейтеромицетов – паразитов растений и представляют тесно скученные многочисленные конидиеносцы, сначала развивающиеся в ткани хозяина под кутикулой или эпидермисом, а затем прорывающие их и выступающие наружу (рис. 228).
Наиболее сложные конидиальные структуры – пикниды. Они имеют шаровидную или кувшиновидную форму, одеты плотной светлой или темной оболочкой с узким отверстием (порусом) на вершине. Внутри пикниды образуются плотным слоем короткие конидиеносцы, на которых образуются конидии, выходящие затем из пикниды через порус или трещины в оболочке, часто в массе слизи (рис. 228). Подробно строение пикнид описано в разделе «Порядок сферопсидные».

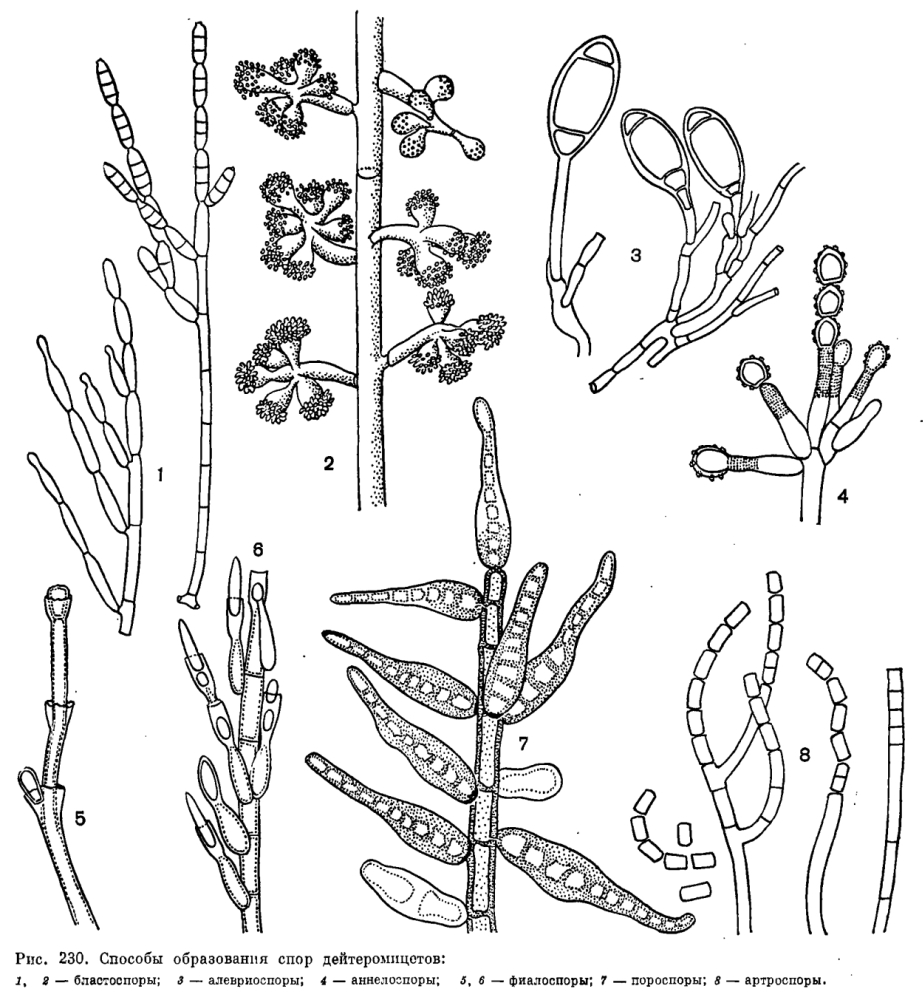
Конидии дейтеромицетов разнообразны по строению. Они могут быть одноклеточными или с различным числом перегородок, иногда муральными (рис. 229). Обычно они шаровидные или эллипсоидные, но у некоторых дейтеромицетов известны нитевидные, звездчатые или спирально закрученные конидии (рис. 229). У водных гифомицетов они обычно три- или тетрарадиальные, реже сигмоидные (рис. 245). Окраска конидий светлая или темная, буроватокоричневая, вследствие присутствия меланинов.

По характеру развития конидии дейтеромицетов относятся к двум группам – талломной и бластической. Талломные конидии (артроспоры и алевриоспоры) развиваются в результате превращения элемента мицелия. Увеличение в размерах и дифференциация таких конидий происходит после их отделения перегородкой (септой) от конидиогенной клетки; таким образом, конидии этого типа происходят из целой клетки. Артроспоры образуются в результате фрагментации конидиеносца или гифы (Geotrichum, Oidiodendron, рис. 230).
Алевриоспоры образуются из отделившейся перегородкой части конидиогенной клетки, посла разрастания превращающейся в зрелую конидию. Они образуются одиночно на вершине ветвей конидиеносца или гиф и часто имеют крупные размеры и утолщенную клеточную стенку (рис. 230). Иногда они развиваются в коротких цепочках. К этому же типу конидий относятся и аннелоконидии или аннелоспоры, часто образующие довольно длинные цепочки. Первая конидия такой цепочки образуется как концевая алевриоспора, затем конидиеносец прорастает через рубец от споры и конец его превращается в следующую алевриоспору. При многократном прорастании конидиеносца через рубцы спор образуется цепочка алевриоспор. Конидиогенная клетка конидиеносца в результате этого процесса удлиняется, и рубцы, остающиеся после отделения алевриоспор, заметны на ней в виде колец. Конидиогенные клетки такого типа названы аннелофорами, а сами конидии – аннелоконидиями. Они известны у многих грибов, например у аннелофоры африканской (Annelophora africana, или скопуляриопсиса короткостеблевого (Scopulariopsis brevicaulis, рис. 230).
Бластический способ образования конидий характеризуется заметным увеличением в размерах зачатка конидии до его отделения перегородкой от конидиогенной клетки. Конидии здесь развиваются таким образом из части клетки, в отличие от конидий первого типа. На основе участия стенки конидиогенной клет ки в формировании стенки конидии разграничивают три типа бластогенных конидий.
У бластоспор, или бластоконидий, все слои стенки конидиогенной клетки участвуют в образовании степки конидии. Бластоспора развивается как вздутие конца конидиогенной клетки, затем отшнуровывающееся от нее и отделяющееся перегородкой. Этот процесс напомипаот почкование у дрожжей. Бластоспоры образуются в акропетальных цепочках (например, у родов Cladosporium, Septonema, рис. 230) или одиночно, часто на небольших зубчиках (например, у рода Artb.rys, рис. 246).
, 
В формировании клеточной стенки пороспор, или пороконидий, принимают участие только внутренние слои клеточной стенки конидиогенной клетки. Они образуются путем почкования через поры в стенках конидиеносцев. Пороспоры обычно толстостенные, развиваются по одной на вершине и по бокам конидиеносца. Конидии этого типа образуются у представителей рода гельминтоспорий (Helminthosporium), а также у некоторых других гифомицетов (рис. 230).
При образовании третьего типа бластогенных конидий – фиалоспор, или фиалоконидий,- клеточная стенка конидии формируется заново, стенка конидиогенной клетки не участвует в ее образовании. Фиалоспоры образуются на фиалидах – конидиогенных клетках, обычно утолщенных у основания и слегка оттянутых в верхней части, у многих грибов с ясно заметным воротничком (например, у родов Phialophora, Catenularia, рис. 230). У других грибов воротничок фиалиды заметен мало или вообще отсутствует, как у пенициллов и аспергиллов (рис. 231).
, 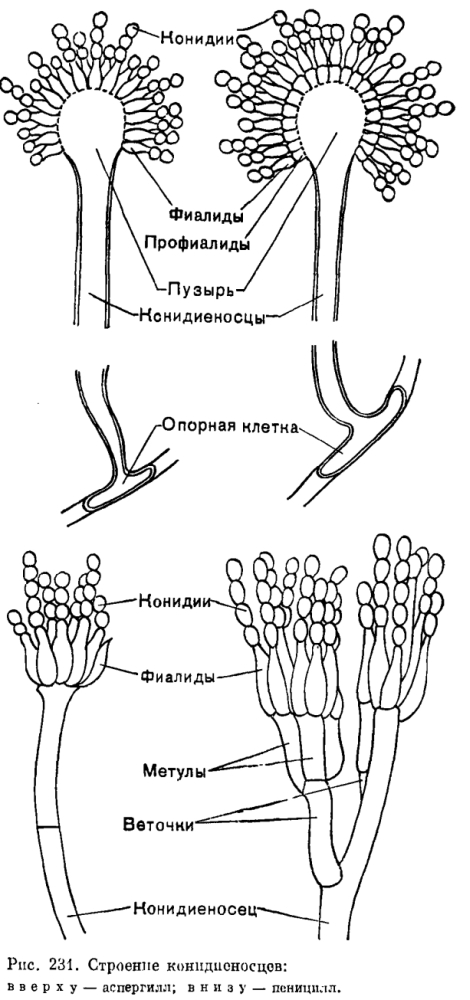
У несовершенных грибов с разными типами конидий хорошо прослеживается параллелизм в усложнении структуры конидиеносцев и их аггрегации, ведущих к увеличению количества спор. Например, среди грибов с фиалоспорами можно обнаружить виды с простыми конидиеносцами (Acremonium), обильно ветвящимися конидиеносцами (пенициллы), коремиями (Stilbella), спородохиями (Tubercularia), ложами (Colletotrichum, МугоШесшт),.пикнидами (Cytospora).
Параллельный ряд усложняющихся форм конидиального аппарата представляют грибы с алевриоспорами. У грибов с пороспорами и артроспорами известны формы только с одиночными конидиеносцами.
Конидии несовершенных грибов, имеющие сухую поверхность, распространяются обычно воздушными течениями. В воздухе можно обнаружить в больших количествах споры грибов из родов алътернария (Alternaria), кладоспорий (Cladosporium), пенициллов и многих других. Если чашку Петри с питательной средой хотя бы ненадолго оставить открытой, в ней уже через 3-4 дня появляются многочисленные колонии этих грибов. Их можно обнаружить даже на высоте более 7000 м над землей. У представителей несовершенных грибов, споры которых погружены в слизь (Acremonium, Verticillium, Graphium), в распространении их участвует вода (часто капли дождя) или мелкие животные (например, жуки-короеды, переносящие конидии графиума – конидиальной стадии возбудителя голландской болезни вязов). При низкой влажности воздуха и высыхании слизи такие споры переносятся также токами воздуха.
Освобождение конидий у несовершенных грибов обычно происходит пассивно. Активное их освобождение наблюдают очень редко. У грибов из родов дейгтониелла (Deightoniella) и курвулярия (Curvularia) конидиогенная клетка теряет воду в результате испарения, в ней развивается отрицательное давление и стенки клетки искривляются. Если процесс потери воды продолжается, оставшиеся ее молекулы сцепляются между собой и со стенками клетки. Вода в клетке находится поэтому в состоянии повышенного поверхностного натяжения, пока не достигается точка, в которой оно снимается путем перехода воды в газообразную фазу. При этом втянутая клеточная стенка конидиогенной клетки внезапно расправляется и сообщает толчок соседней с ней клетке – конидии, которая отбрасывается в воздух (Д. С. Мередит).
Размножение несовершенных грибов происходит только бесполым путем, поэтому можно было бы предположить, что все особи, происшедшие из конидий, образовавшихся из одной колонии, и последующие их поколения будут развиваться практически в виде чистых линий и изменяться очень мало, только за счет немногочисленных спонтанных мутаций. Но в природе у несовершенных грибов этого никогда не наблюдается. Они представляют, напротив, одну из наиболее вариабильных и подвижных в экологическом отношении групп грибов, именно поэтому весьма широко распространенную в природе и заселяющую самые разнообразные субстраты. Это объясняется тем, что у несовершенных грибов очень часто мицелий гетерокариотичен, т. е. содержит генетически различные ядра.
Гетерокариоз известен в различных таксономических группах грибов – у оомицетов, аскомицетов, зигомицетов, но у несовершенных грибов он представляет собой основной механизм изменчивости. В процессе развития гетерокариотического мицелия число ядер того или иного типа может варьировать в зависимости от изменения условий среды, обеспечивая таким образом адаптацию гриба к этим изменениям. Так, пеницилл распростертый (Penicillium expansum) в природе всегда встречается в гетерокариотическом состоянии. Его гомокариотические изоляты (с генетически однородными ядрами) обычно растут хуже, чем исходный гетерокарионт. Содержание ядер разных генетических типов у этого гриба регулируется условиями среды, например составом питательных сред: при изменении содержания в среде яблочной пульпы число ядер одного из генетических типов (А) колеблется от 13 до 58%. Такие же колебания числа ядер разных типов в зависимости от условий обитания обнаружены и у некоторых фузариумов.
Образование гетерокариотического мицелия у несовершенных грибов может происходить несколькими путями. У одних из них на гетерокариотическом мицелии образуются конидии, содержащие генетически различные ядра. Из таких конидий снова развивается гетерокариотический мицелий. У аспергилла угольного (Aspergillus carbonarius), например, конидии содержат по 2-5 ядер. Гетерокарионты между диким типом с черными конидиями и мутантом со светло-коричневыми конидиями образуют споры промежуточной окраски, которые, в свою очередь, дают колонии также смешанного типа. Если конидии на гетерокариотическом мицелии образуются из одноядерных конидиогенных клеток, они гомокариотичны, т. е. содержат ядра только одного генетического типа. У другого вида аспергиллов – A. versicolor – гетерокарионты между диким штаммом с зелеными конидиями и мутантом с белыми конидиями образуют головки с цепочками белых и зеленых конидий. Из таких конидий вырастает гомокариотический мицелий. Гетерокариотичность у него может восстанавливаться в результате образования анастомозов между гифами с генетически различными ядрами, а также в результате мутаций в отдельных ядрах мицелия.
В некоторых случаях гетерокариоз может быть основой не только изменчивости и адаптации гриба в результате изменения в его мицелии числа генетически различных ядер, но и рекомбинации признаков. Однако рекомбинация у несовершенных грибов происходит не при мейозе, а при митозе в изредка образующихся в гетерокариотическом мицелии диплоидных ядрах. Такие ядра могут гаплоидизироваться в результате утраты ими хромосом.
Процесс рекомбинации такого типа был назван парасексуальным процессом (Дж. Понтекорво). Он слагается из нескольких этапов: слияния ядер гетерокариона и образования диплоидного гетерозиготного ядра; размножения таких гетерозиготных ядер в гетерокариотическом мицелии вместе с гаплоидными; митотической рекомбинации (при митозе) во время размножения диплоидных ядер; вегетативной гаплоидизации диплоидных ядер в результате утраты ими хромосом. Парасексуальный процесс известен сейчас у многих несовершенных грибов, однако его наблюдают у них только в условиях лаборатории; какова же его роль в естественных условиях обитания, пока не установлено.
Несовершенные грибы широко распространены в природе во всех районах земного шара. Многие представители этой группы обитают как сапрофиты в почве, составляя большую часть почвенных грибов. Они в изобилии встречаются на различных растительных субстратах, реже на субстратах животного происхождения. Эти группы несовершенных грибов принимают активное участие в разложении органических остатков и в почвообразовательном процессе. Некоторые сапрофитные несовершенные грибы вызывают плесневение пищевых продуктов и различных промышленных изделий.
Многочисленна группа несовершенных грибов, паразитирующих на высших растениях. Они вызывают многочисленные серьезные болезни сельскохозяйственных культур, приносящие большой экономический ущерб. Некоторые представители этого класса вызывают заболевания у животных и человека. Развиваясь на зерне и других продуктах питания, отдельные несовершенные грибы выделяют в них токсины, которые могут вызвать тяжелые отравления при использовании таких продуктов в пищу человеком или кормлении ими животных.
Среди несовершенных грибов известны многочисленные продуценты биологически активных веществ, используемые при производстве антибиотиков (пенициллина, гризеофульвина, фумагиллина, трихотецина), различных ферментов и органических кислот. Несовершенные грибы, паразитирующие на насекомых-вредителях и грибах, патогенных для растений, а также хищные грибы, уничтожающие фитонематод, используют для разработки биологических методов защиты растений от вредителей и болезней.
Широко используемая сейчас всеми микологами система несовершенных грибов, разработанная П. А. Саккардо в 1880 г., заведомо искусственна и служит только более или менее удобной схемой для их упорядочения и определения. Несовершенные грибы подразделяются в ней на три порядка на основе строения конидиального аппарата. Порядок гифомицеты (Hyphomycetales) объединяет несовершенные грибы, образующие одиночные конидиеносцы или конидиеносцы, соединенные в коремии и спородохий. К порядку меланкониевых (Melanconiales) относятся несовершенные грибы, образующие ложа, а к порядку сферопсидных (Sphaeropsidales) – образующие пикниды.
Некоторые микологи (С. Дж. Xьюз, К. В. Субраманиан, К. Тубаки, С. Нильссон, Дж. Л. Баррон и другие) делят несовершенные грибы на разное число групп по типу спорогенеза, основными из которых являются: бластоспоровые (Blastosporae), алевриоспоровые (Aleuriosporae), фиалоспоровые (Phialosporae), пороспоровые (Рогоsporae) и артроспоровые (Arthrosporae).
Однако новая система несовершенных грибов находится пока в процессе разработки, она охватывает далеко не все известные сейчас несовершенные грибы. Морфогенез конидий хорошо изучен только у гифомицетов, но он известен лишь у отдельных представителей двух других порядков.
В настоящей книге мы пользуемся системой П. А. Саккардо, охватывающей все многообразие несовершенных грибов.
Жизнь растений: в 6-ти томах. – М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
Источник